Verbreitung und Ökologie der Kriebelmacken (Diptera, Simuliidae) in der Schweiz Von ROLF GLATTHAAR. ZÜRICH Inhaltsverzeichnis 1. Einleitung 71 2. Untersuchungsgebiet, Material und Methoden 72 a) Aufsammeln der Kriebelmücken 72 b) Chemische Analysen 81 3. Verbreitung der Arten in der Schweiz 82 4. Phänologie der Simuliidae 93 5. Assoziationen zwischen Simuliidenarten 96 6. Autökolo g ie der präimaginalen Kriebelmücken 104 7. Prognostische Bedeutung der Analysen 114 S. Diskusslon I16 9. Zusammenfassung — Resume — Summary 119 10. Literaturverzeichnis 121 1. Einleitung Die Kriebelmücken (Simuliidae), eine der individuenreichsten Dipteren-Familien, kommen mit Ausnahme des vergletscherten Hochgebir g es in allen Gegenden der Schweiz vor. Ihre Larven und Puppen leben im Benthos^fast aller Schweizer Fliess- gewässer. Die Weibchen saugen Blut an Mensch und Tier. In einigen Gebieten treten sie besonders im Frühjahr so zahlreich auf, dass sie dem Vieh durch ihre Stiche gefährlich werden können (BARANOV, 1939; WILHELMI, 1920; RUEHM, 1967; ENIGK, 1955). In der Schweiz kam es erstmals 1968 zu Todesfällen, als deren Ursache Kriebel- mückenstiche nach g ewiesen wurden (EcKERT et al., 1969). Trotz der starken Präsenz der Kriebelmücken liegt mit Ausnahme eini ger lokaler Nachweise (GALLI-VALERIO, 1927; KUHN, 1952; ECKERT et a l. , 1969; L. DAVIES, 1957; D. M. DAVIES, 1961: ZWICK, 1974) noch keine Darstellung ihrer g eographischen Verbreitung oder der
Transcript
Verbreitung und Ökologie der Kriebelmacken(Diptera, Simuliidae) in der Schweiz
Von
ROLF GLATTHAAR. ZÜRICH
Inhaltsverzeichnis
1. Einleitung 71
2. Untersuchungsgebiet, Material und Methoden 72a) Aufsammeln der Kriebelmücken 72b) Chemische Analysen 81
3. Verbreitung der Arten in der Schweiz 82
4. Phänologie der Simuliidae 93
5. Assoziationen zwischen Simuliidenarten 96
6. Autökologie der präimaginalen Kriebelmücken 104
7. Prognostische Bedeutung der Analysen 114
S. Diskusslon I16
9. Zusammenfassung — Resume — Summary 119
10. Literaturverzeichnis
121
1. Einleitung
Die Kriebelmücken (Simuliidae), eine der individuenreichsten Dipteren-Familien,kommen mit Ausnahme des vergletscherten Hochgebir ges in allen Gegenden derSchweiz vor. Ihre Larven und Puppen leben im Benthos^fast aller Schweizer Fliess-gewässer. Die Weibchen saugen Blut an Mensch und Tier. In einigen Gebieten tretensie besonders im Frühjahr so zahlreich auf, dass sie dem Vieh durch ihre Stichegefährlich werden können (BARANOV, 1939; WILHELMI, 1920; RUEHM, 1967; ENIGK,
1955). In der Schweiz kam es erstmals 1968 zu Todesfällen, als deren Ursache Kriebel-mückenstiche nach gewiesen wurden (EcKERT et al., 1969). Trotz der starken Präsenzder Kriebelmücken liegt mit Ausnahme eini ger lokaler Nachweise (GALLI-VALERIO,1927; KUHN, 1952; ECKERT et a l. , 1969; L. DAVIES, 1957; D. M. DAVIES, 1961:ZWICK, 1974) noch keine Darstellung ihrer geographischen Verbreitung oder der
72 Vierteljahrsschrift der Naturforschenden Gesellschaft in Zürich I978
Abundanz einzelner Arten in der Schweiz vor, wie es sie für andere europäischeLänder bereits gibt. Mir bot sich hauptsächlich in den Sommermonaten der Jahre1973 und 1974 Gelegenheit, dieser Fra ge nachzu gehen. Ausserdem untersuchte icheine Anzahl von Umweltfaktoren, welche die häufigeren Arten beeinflussen könnten.
Bei der Bestimmung, Taxonomie und Nomenklatur stützte ich mich auf die Arbei-ten von KNOZ (1965), ZWICK (1974), RIVOSECCHI und CARDINALI (1975) sowie Rus-zow (1959-1964), auf die für die Nachweise ausserschweizerischer Funde sowie fürdie ältere Literatur verwiesen wird.
Dank. Herr Prof. Dr. J. ECKERT, Direktor des Institutes für Parasitologie derUniversität Zürich, schlug das Thema vor. Herr Prof. Dr. W. RUEHM, Institut fürParasitolo gie der Tierärztlichen Hochschule in Hannover, jetzt Zoologisches Institutder Universität Hamburg, ermö glichte mir einen einmonati gen Studienaufenthalt inHannover und vermittelte mir die Kenntnis der mitteleuropäischen Simuliidenarten.Ausserdem überliess er mir eine Serie individuell aufgezo gener Odagmia-ornata-Ima-gines für eine numerisch-taxonomische Analyse. Frau Dr. H. ZWICK-PODSZUHN, Max-Planck-Institut für Limnologie in Schlitz (Hessen, BRD), stand mir in taxonomischenFragen zur Seite und verifizierte mehrere Kriebelmückenproben. Herr Prof. Dr. H. BUR-
LA, Direktor des Zoologischen Museums der Universität Zürich, und Oberassistent Dr.G. BAECFLI gaben Ratschläge und halfen bei der Abfassung des Manuskripts. AssistentDipl.-Math. W. STAHEL beriet mich in Fragen der numerischen Taxonomie und inder rechnerischen Auswertun g, die ich am Rechenzentrum der Universität durch-führte. Herr Prof. Dr. W. STUMM, Direktor der Eid genössischen Anstalt für Wasser-versorgung, Abwasserreini gung und Gewässerschutz (EAWAG) in Dübendorf ZH,stellte die hydrochemischen Daten der Lan gzeitnntersuchun g an der Glatt und amChriesbach zur Verfü gung und erlaubte mir, unter der Anleitung von Mitarbeiternder Chemischen Abteilun g, Wasseranalysen an der EAWAG durchzuführen. HerrProf. Dr. H. AMBUEHL, Leiter der Limnologischen Abteilun g der EAWAG, überliessmir die Daten der Querschnittsanalyse der schweizerischen Fliessgewässer des Jahres1974 (Projekt Mapos), Herr Dr. P. PERRET und Herr F. STOESSEL die Ergebnisse derFaunenanalysen, die von verschiedenen Mitarbeitern der Limnologischen Abteilungdurchgeführt worden waren. Herr Dr. J. ZOBRIST, Chemische Abteilung der EAWAG,beriet mich in hydrochemischen Fragen und be gutachtete das Manuskript. DieKARL-HESCHELER- Stiftung gewährte einen Kostenbeitra g an die Sammelexkursionen,für welche meine Eltern ihren Wa gen zur Verfü gung stellten.
Die Naturforschende Gesellschaft in Zürich leistete einen Beitrag an die Druck-kosten.
2. Untersuchungsgebiet, Material Hnd Methoden
a) Aufsammeln der Kriebelmiicken
Vom April 1973 bis Ende Oktober 1974 suchte ich etwa 250 Probestellen auf, diesich mit Ausnahme der Kantone Appenzell und Genf über die ganze Schweiz ver-teilen. Sie umfassen alle in der Schweiz vorkommenden Typen von Fliessgewässern.
Jahrgang 123 R. GLATTHAAR. Verbreitung und Ökologie der Kriebelmücken 73
Die Sammeltätigkeit gliederte sich in zwei Serien. Vom Frühjahr 1973 bis zumFrühlin g 1974 bearbeitete ich fast alle grösseren Wasserläufe des Kantons Zürich,von den zahlreichen Wald- und Wiesenbächen eine Auswahl. 23 Fundstellen, diereichlich Simuliiden lieferten, sowie zwei Orte, die zu keiner Jahreszeit von ihnenbesiedelt waren, unterzo g ich einer Lan gzeitbeobachtung. In der Wachstumssaison,Anfang April bis Ende Oktober, nahm ich alle drei bis fünf Wochen, in der folgendenkalten Jahreszeit alle sechs bis acht Wochen je eine Kriebelmücken- und eine Wasser-probe. Mit wöchentlichen Stichproben führte die Eid genössische Anstalt für Wasser-versorgung. Abwasserreinigung und Gewässerschutz (EAWAG) in der Glatt und imChriesbach ihre Langzeitanalysen durch (ZoBRIST et al., 1976), deren Daten ich miteinbezog.
Im Sommer 1974 nahm die EAWAG eine Untersuchung über den Zustand derschweizerischen Fliess gewässer in An griff (PERRET, 1977), in deren Rahmen alle grös-seren Wasserläufe mit mindestens einer Probestelle berücksichti gt wurden, an welcherje eine bis drei Wasserproben und eine Faunenprobe genommen wurden. Es handeltsich somit um eine «Momentaufnahme», nachstehend «Querschnittsuntersuchung»genannt. Aus dieser wählte ich hundert Probestellen aus, die sich über das ganzeAlpengebiet sowie die Voralpen und das Flachland der Kantone Aargau, Bern, Luzernund Schwyz verteilen. An 85 Stellen wurden Kriebelmücken gefunden.
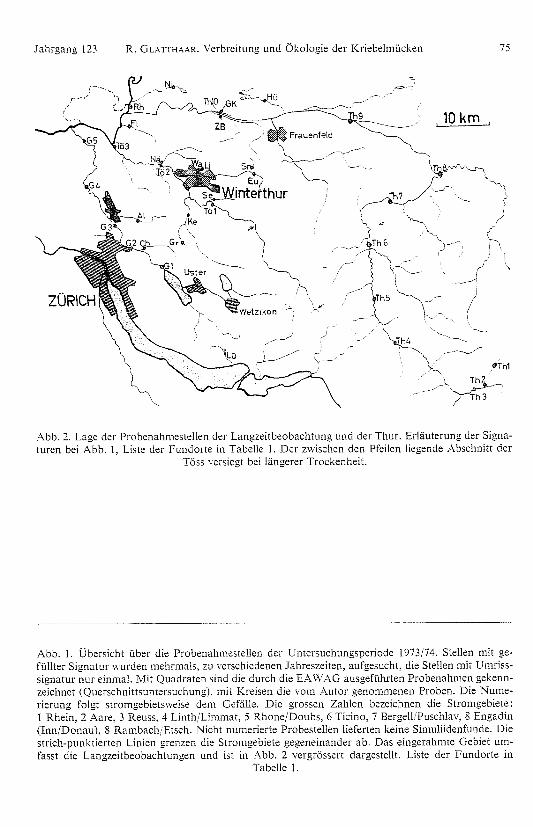
Die Abbildun gen 1 und 2 sowie Tabelle 1 geben eine Übersicht über die Fundorte.Die auf der Schweizerkarte (Abb. 1) ein getragenen werden im fol genden mit einerdreistelligen Zahl zitiert, deren erste Ziffer die Strom gebietszugehöri gkeit angibt(grosse Zahlen in Abb. 1), die beiden fol genden in der Reihenfolge abnehmenderHöhenla ge die Nummer der Probestellen (kleine Zahlen). Für die Langzeitunter-suchun g sowie für die Thur wählte ich als Code die Abkürzung des Gewässernamensund numerierte, wo nötig. die Probestellen in der Fliessrichtung.
Tabelle 1. Liste der Probenahmestellen der Langzeit- und der Querschnlttsuntersuchung. Codes wiein Abb. 1 und 2. In Klammern steht die Anzahl der entnommenen Kriebelmücken- und Wasser-proben; fehlende Angabe bedeutet 1 Probe. Die Stellen der Langzeituntersuchung sind mit einem
Stern (") markiert.
Code Gewässer, Ort
101 Vorderrhein unterhalb Disentis102 Glogn bei Ilanz103 Hinterrhein bei Splügen104 Averserrhein unterhalb Ausserferrera105 Hinterrheln unterhalb Thusis106a Albula bei Fürstenaubruck106b Seitenbach unterhalb Fürstenaubruck107 Sertigbach oberhalb Davos108 Dischmabach oberhalb Davos109 Plessur bei Molins110 Landquart bei Auja oberhalb Klosters (2)111 Landquart unterhalb Klosters (2)112 Schlappina, Alp Schlappin oberhalb Klosters113 Landquart bei Küblis (2)114 Rhein bei der Brücke östlich Haag
74 Vierteljahrsschrift der Naturforschenden Gesellschaft in Zürich 1978
Jahrgan g 123 R. GLATTHAAR. Verbreitung und Ökolo gie der Kriebelmücken 75
Abb. 2. Laoc der rmucouxmoncteoder LangzeitbeobachtungunducrTuucsuämtemogm,Siooutureo bei Abb. l, Liste der Fundorte in Tabelle l. Der zniscu o den pfeo n lie gende Abschnitt der
Abb.|.Ubecsicb,u berdie9mhcnahmcorlleodc,linteoucouwppcdodem73/74.8tellenoit:e-[n8terSienatu,vurucooebonvb.zuw,ozhiedcneo/axrmziteo.uufgcsucht.uiea^elleomhDmrss-sigomurourriomal.aait Quadraten sind die durch die EAWAG ausgeführten Probenahmen gekenn-zeichnet (Querschnittsuntersuchung), mit Kreisen die vom Autor genommenen Proben. Die Nume'rieruon[u|»cstmmoruict,nciseucmorfäDc.oieomoeozumcourzcicbocodieStmmgehiete:lnhdn.zAvre.smeuo.4Dn,x/Limmocsmbuoc/oouhs.oTiciou.7a,rgcU/puscN^xoEoouuio(/oo/oovou). 8 Rambach/Etsch. Nicht numerierte Probestellen lieferten keine Simuüid^ob^ouo. Die,t,ich-puoxhrrteoLinicugre000uiestmm:ebietogeocociouoacruhomringec`bmtco,bictunu'5uut die Lan gzeitbeobachtun gen und ist in Abb. 2 ver grössert dargestellt. Liste der Fundorte in
Tabelle I.
76 Vierteljahrsschrift der Naturforschenden Gesellschaft in Zürich 1978
Code Gewässer, Ort
115 Simmibach unterhalb GaIns116 Rheintaler Binnenkanal östlich Sevelen117 Rhein bei Wagenhausen118 Biber unterhalb Ramsen119a Rhein bei Rheinklingen119 b Geisslibach unterhalb Diessenhofen120a Rhein beim Kloster St. Katharinenthal120b Seitenbach bei Paradies121 Rhein bei Zurzach122 Sissle oberhalb Sisseln123 Möhlinbach oberhalb Möhlin124 Ergolz oberhalb Augst125 St.-Alban-Teich, Baselstadt126 Birs bei Basel127 Lucelle-Teichausfluss128 Etang de Bollement, Auslauf129 Scheltenbach unterhalb Schelten
201 Lombach unterhalb Habkern202 Aare bei Münsingen203 Aare, Hagneckkanal oberhalb Hagneck204 Kleine Simme oberhalb Zweisimmen205 Simme bei Oey206 Chirel bei Oey207 Gürbe bei Belp208 Saane bei Saanen209 Sarine (Saane) bei La Tine210 La Gerine bei Marly-le-Grand211 Broye unterhalb Payerne212 Emme bei Burgdorf213 Emme unterhalb Luterbach214 Quell- und Waldbäche, Balmberg oberhalb Günsberg (2)215 Dünnern bei Laupersdorf216 Dünnern bei Niederbuchsiten217 Aare bei Wolfwil218 Köllikerbach unterhalb Kölliken219 Napfgebiet, Waldbäche220 Wigger oberhalb Willisau221 Mauensee-Ausfluss222 Wigger bei Brittnau223 tirke bei Oberentfelden224 Wyna unterhatb Zetzwil225 Wyna bei Suhr226 Suhre unterhalb Schöftland227 Suhre unterhalb Suhr228 Ron unterhalb Hochdorf229 Aabach bei Mosen (2)230 Aabach unterhalb Seon231 Aabach bei Wildegg232 Bünz unterhalb Bünzen233 Bünz unterhalb Dottikon234 Bünz bei Wildegg235 Aare bei Wildegg
Jahrgang 123 R. GLATTHAAR. Verbreitung und Ökologie der Kriebelmücken 77
Code Gewässer , Ort
301 Reuss bei Andermatt302 Reuss bei Erstfeld303 Schächenbach bei Unterschächen (2)304 Isleterbach bei Isleten305 Muota unterhalb Muotathal306 Steiner Aa oberhalb Steinen307 Steiner Aa unterhalb Steinen308 Rigi-Aa bei Arth309 Engelberger Aa bei Engelber g, Titlisbahnstation310 Engelberger Aa bei Büren NW311 Melch-Aa oberhalb Sarnen312 Sarner Aa bei Alpnach313 Waldemme bei Flühli314 Kleine Emme bel Ebnet315 Fontannen bei Doppleschwand316 Kleine Emme bei Maltee (3)317 Ränggbach oberhalb Blatten LU318 Kleine Emme bei Emmenbrücke (2)319 Rotbach bei Inwil320 Reuss oberhalb Hellingen321 Lorze, A gerisee-Ausfluss bei Unterägeri322 Lorze bei Baar (2)323 Lorze, Zugersee-Ausfluss bei Cham324 Jonen unterhalb Jonen
401 Fätschbach, Urnerboden (4)402 Linth bei Hasten403 Linth, Walensee-Einmündung404 Seez bei Flums405 Minster bei Atmig406a Sihl bei Studen SZ (2)406b Waag bei Studen SZ (2)407 Alp bei Trachslau (2)408 Alp bei Biberbrugg (2)409 Biber, Altmatt410 Biber bei Biberbrugg (2)411 Wiesenbach bei Egg bei Einsiedeln412 Sihl bei Sihlbrugg (4)413 Sihl bei Langnau am Albis (2)414 Sihl beim Zürcher Hauptbahnhof415 Limmat beim Kloster Fahr (2)416 Furtbach bei Würenlos417 Reppisch bei Landikon418 Reppisch bei Bergdietikon419 Reppisch unterhalb Dietikon420a Hüttenersee-Auslauf oberhalb der Schleuse (2)420b Hüttenersee-Ausfluss unterhalb der Schleuse (2)
501 Saltina oberhalb Brig502 Turtmänna bei Turtmann503 a La Printse unterhalb Beuson503b La Borgne bei Bramois504 La Salentse bei Saillon505 La Vizze bei Monthey
78 Vierteljahrsschrift der Naturforsch:nden Gesellschaft in Zürich 1978
Code Gewässer, Ort
506a La Gryonne oberhalb Bex506b L'Avangon oberhalb Bex507 Grande Eau oberhalb Aigle508 Eau Froide oberhalb Villeneuve509 Doubs bei St-Ursanne510 L'Atlaine oberhalb Alle
601 a Ritomsee-Zuflüsse, Val Piora601 b Ticino oberhalb Airolo602a Ticino bei Faido (2)602b Seitenbach bei Quinto603 Ticino unterhalb Giornico (4)604 Ticino bei Claro (2)605 Brenno bei Campo Blenio606a Brenno unterhalb Acquarossa (3)606b Quellbächtein am Pizzo Molare oberhalb Acquarossa606c Bächlein bei der Capanna Adula CAS607 Calancasca im Calancatal608 Moesa unterhalb Mesocco (3)609 Moesa bei Grono (2)610 Morobbia unterhalb Giubiasco611 Sementina unterhalb Sementina612 Magadinoebene, linker Seitenkanal613 Verzasca bei Lavertezzo614 Maggia bei Ponte Brolla615 Isorno bei Intraena616 Bach bei Bosco{Gurin617 Vedeggio bei Vira (2)618 Cassarate bei Canobbio619 Vedeggio bei Agno620 Magliasina bei Brenno621 Magliasina unterhalb Magliaso622 Tresa bei Ponte Cremenaga (2)623 Mara bei Maroggia624 Breggia unterhalb Chiasso625 Doveria unterhalb Ruden/condo
701 MaiIa unterhalb Casaccia702 Maira bei Castasegna703 Mera bei Piuro/Borgonovo, Italien704 Poschiavino bei Le Prese705 Poschiavino unterhalb Brusio706 Quellbäche Alp Grüm / Alpe Palü
801 Inn beim \Ialoja-Hotel802 Wiesenbächlein auf der Maloja-Passhöhe803 a Fedacla unterhalb Sils-Maria803 b Silsersee-Ausfluss (so genannter See-Inn) (2)804a Auslauf des Lej Nair (3)804b Hahnensee-Auslauf805 Quellbach oberhalb Hotel St. Moritzbad (3)806 Inn bei Samedan (3)
901 Rambach (II Rom) unterhalb Müstair
Jahrgang 123 R. GL 1-rHAAR. Verbreitung und Ökolo g ie der Kriebelmücken 79
Code Gewässer, Ort
Al *Altbach unterhalb Bassersdorf (9)Ch * Chriesbach bei der EAWAG (18)Eu * Eulach bei der Kläranlage Elgg (8)Fl * Flaacherbach unterhalb Flaach (10)
G 1 *Glatt bei Schwerzenbach (14)G 2 *Glatt bei Opfikon (7)G 3 * Glatt bei Rümlang (6)G 4 * Glatt bei Niederglatt (7)G 5 * Glatt bei Glattfelden (16)
Hü Hüttwilerseebach (5, ohne Wasser)Ke * Kempt bei der Kläranlage Illnau-Effretikon (11)Li Lindbergwaldbäche (8)Lü * Lützelseeausfluss (6)Nä * Näfbach bei Pfungen (11)
Ni Niderbach bei Dachsenhausen (6)Gr * Grendelbach oberhatb Bisikon (10)Rh *Rhein bei Ellikon am Rhein (10)Se * Sennhofbach bei Sennhof im Tösstal (6)Sn * Schneitbach unterhalb Schneit/Hagenbuch (5)
Th 1 Säntisthur, Thurboden (3, ohne Wasser)Th 2 Säntisthur bei Unterwasser (3, ohne Wasser)Th 3 Thur bei Alt St. Johann (4, ohne Wasser)Th 4 Thur bei Nesslau (4, ohne Wasser)Th 5 Thur unterhalb Ebnat-Kappel (4, ohne Wasser)Th 6 Thur unterhalb Bütschwil (4, ohne Wasser)Th 7 Thur oberhalb Uzwil (4. ohne Wasser)Th 8 Thur bei Bischofszell (3, ohne Wasser)Th 9 Thur bei Amlikon (3, ohne Wasser)Th 10 * Thur bei Altikon (10)
GK *rechtsseitiger Grundwasserkanal bei Th 10 (8)ZB *sogenannter Zürcher Binnenkanal links der Thur bei Th 10 (9)
Tö 1 * Töss bei Sennhof (10)Tö 2 ;Töss bei Pfun gen (7)Tö 3 . Töss oberhalb Tössegg, Rheinmündung (10)
Wa * Walcheweiher-Auslauf im Lindbergwald (6)
Bei jeder Probestelle beider Serien protokollierte ich die folgenden Umwelt-faktoren:
— Höhe über Meer.
— Wassertemperatur.
— Flussbreite.
Wassertiefe, an drei Stellen des Flussquerschnittes.
— Fliess geschwindi gkeit, an denselben drei Stellen, mit Messflügel geräten der FirmenSTOPPANI AG, Bern. und E. SCHILTKNECHT AG , Gossau ZH.
— Abflussmen ge, errechnet aus den Breiten-, Tiefen- und Fliessgeschwindigkeitsmes-
80 Vierteljahrsschrift der Naturforschenden Gesellschaft in Zürich 1978
sungen. Für den Rhein bei Ellikon (Rh in Abb. 2) griff ich auf die Daten desElektrizitätswerkes der Stadt Schaffhausen zurück (DEMMERLE, 1966).
— Turbulenz am Entnahmeort der Kriebelmücken, anhand folgender subjektiv an-wendbaren Skala:1 Glatte, stellenweise leicht gekräuselte Wasseroberfläche, geräuschloser Fluss,
von Makrophyten bewachsen. Bachbett meist sandig oder schlickig. Meist beiSeeausläufen.
2 Wasseroberfläche gekräuselt, murmelndes Fliess geräusch. Einzelne Strom-schnellen, Bachbett mit Grobkies. Wenige Makrophyten.
3 Rauschender Fluss mit zahlreichen Stromschnellen und kleineren Stufen. Bach-bett mit Geröll.
4 Starkes, bei grösseren Gewässern donnerndes Rauschen, grössere Schnellen undStufen. Felsblöcke im Bachbett. Ausbildung einer Spritzzone.
— Hydrophytenbewuchs des Flussbettes, als relativer Anteil aller subaquatisch wur-zelnden Makrophyten plus der ins Wasser ra genden Teile der Uferpflanzen an derWasseroberfläche.
— Grad der Uferbeschattung, aus gedrückt durch den prozentualen Anteil der Him-melshemisphäre, der durch Pflanzen, Bauten oder Gebir ge ständi g verdeckt ist.
— Mittlere Grösse der Substrateinheiten (Länge von Blättern und Aster': grössterDurchmesser von Steinen, Plastikfolien usw.).
— Oberfläche des nach Kriebelmücken ab gesuchten Substrates, unter Ausschlussnicht besiedelbarer Teile wie etwa die Auflageflächen von Steinen.
Bei kleinen Proben oder geringer Populationsdichte las ich die Tiere mit einemPinsel oder einer Pinzette vom Substrat ab und fixierte sie mit 70% Alkohol. GrössereProben, vor allem solche, die dicht mit Larven und Puppen besiedelte Pflanzenteileenthielten, transportierte ich in frisch benetzten, nötigenfalls gekühlten Thermos-behältern. Die Gele ge, Larven und Exuvien reini gte ich von Al gen und Detritus undfixierte sie mit 70% Alkohol. Wenn ich für die Artbestimmun g Ima gines benötigte,setzte ich die Puppen in etwa 10 cm lan ge.. abtropfnasse Glastuben zu 50 cm 3 Inhalt.Geeignete Desinfektionsmassnahmen sowie eine sor gfältige Manipulation beugtendem Pilzwachstum vor. Die Tuben mit den einzeln gehaltenen Puppen standen, mitder Öffnun g nach unten, in einem Plastikbecken an einem hellen, kühlen Platz, bisdie Ima gines schlüpften. Mit diesem Verfahren erzielte ich bei genügend reifen PuppenSchlüpferfolge von 60 bis 100%. Nachdem die Ima gines mindestens einen Tag Zeitgehabt hatten, um auszuhärten und auszufärben, fixierte ich sie.
Die für die biozönotische Gliederun g der Thur, Töss und Glatt verwendeten Probenbestimmte und zählte ich vollständi g aus, bei den übri gen Gewässern nur die klei-neren. Bei grösseren wertete ich nur etwa 200 Tiere aus, wobei Altlarven und Puppenden Vorzu g erhielten. Den Rest prüfte ich auf Vertreter seltener Arten und schätztedie Anzahl Individuen je häufige Art.
Um die Phänologie der häufi geren Arten erfassen zu können, untersuchte ich auchdie Altersstruktur in den grösseren Proben. Ich unterschied sechs Altersklassen, diein der warmen Jahreszeit Abschnitten von einer bis zwei Wochen entsprechen:
Jahrgang 123 R. GLATTHAAR. Verbreitung und Ökologie der Kriebelmücken 81
1. Gelege2. Junglarven (erstes bis fünftes Stadium)3. Sechstes Larvenstadium4. Siebentes Larvenstadium5. Puppen6. Exuvien und Kokons (entsprechen der Anzahl Imagines)
Um verschieden grosse Proben miteinander vergleichen zu können, schätzte ichdie Besiedlungsdichte je Art, wobei ich mich einer logarithmischen Skala bediente,deren Einheit einer Zehnerpotenz entspricht:
1 1 bis 10 Individuen pro m''2 10 bis 100 Individuen pro m'3 100 bis 1 000 Individuen pro m24 1 000 bis 10 000 Individuen pro m25 mehr als 10 000 Individuen pro m2
b) Chemische Analysen
Soweit die Wasserproben nicht unmittelbar nach dem Einsammeln analysiert wer-den konnten, wurden sie in Glas- oder imprägnierten Polyäthylenflaschen an einenkühlen Ort verbracht, wo die Temperatur 10° C nicht überstieg. Tabelle 2 gibt Aus-kunft über die erfassten hydrochemischen Faktoren. Die Analysen zur Langzeit-
Tabelle 2. Übersicht über die untersuchten hydrochemischen Faktoren
82 Vierteljahrsschrift der Naturforschenden Gesellschaft in Zürich 1978
beobachtun g führte ich selbst durch, mit Ausnahme der Stellen in der Glatt und imChriesbach; die Querschnittsanalysen besorgte die EAWAG im Sommer 1974. Infolgeder zeitlichen Verschiebung und der etwas abweichenden Zielsetzun g unterscheidensich die Programme in einigen Faktoren. Den Analysen lagen die Vorschriften desSchweizerischen Lebensmittelbuches, Kapitel 27, zugrunde sowie die Methoden inden «Vorläufigen Empfehlungen über die regelmässige Untersuchun g der schwei-zerischen Oberflächengewässer» (Eidgenössisches Departement des Inneren, 29. Mai1974).
3. Verbreitung der Arten in der Schweiz
Im Laufe der Sammeljahre 1973 und 1974 erbeutete ich über 250000 Larven undPuppen sowie etwa 50000 schwärmende Weibchen, unter denen sich auch einigeDutzend Männchen befanden. Die Ima gines fin g ich auf der Viehweide und amweidenden Vieh bei Thalheim an der Thur, wo sie durch ihre Stiche lästi g wurden,sowie am Stadtrand von Winterthur , wo sie mich an fl o gen. Beim Entlan gstreifen ander Ufervegetation von Thur, Töss und Gla tt gingen nur vereinzelte Imagines insNetz. Alle 28 nachstehend aufgeführten Arten sind durch Puppenfunde belegt; vonden meisten züchtete ich aus Puppen insgesamt über 1000 Imagines. Trotz den grossenIndividuenzahlen lassen sich erst im Kanton Zürich Verteilung und Abundanz dereinzelnen Arten detailliert an geben, da die übri gen Gebiete noch zuweni g durchsuchtworden sind. Der gegenwärti ge Stand der Chorologie zei gt, dass keine Art auf dieSchweiz beschränkt ist und dass der Alpenkamm für keine Art eine Verbreitungs-grenze darstellt.
Die folgende Artenliste gibt die Anzahl der gesammelten präimaginalen Individuennnd in der Reihenfolge abnehmender Populationsdichte die Codes der wichtigstenFundorte mit einer kurzen Beschreibung der Fundumstände. Bei den selteneren Artenwerden ausserdem ökolo gische Beobachtun gen mitgeteilt. Die in der Schweiz erstmalsnachgewiesenen Arten sind mit Stern (') vor dem Namen gekennzeichnet.
Unterfamilie Prosimuliinae ROUBAUD 1906
Tribus Gymnopaidini
Genus Twinn a STONE und JAMNB.vCK 1955
Twi,mia hvdroides NOVAK 1956
Fundorte: 401. 805, 706.Etwa 300 Larven und Puppen.
Von der in der Tschechoslowakei entdeckten Art (KNOZ, 1965) gibt es erst dreiaussertschechische Fundorte: Polen. der Schwarzwald und das Oberen gadin, Stelle805 (D. M. DATES 1961). Mitte Juni 1974 und 1977 konnte ich den schweizerischenNachweis bestäti gen. Etwa 15 Minuten oberhalb des St. Moritzer Badhotels am Wegzum Hahnensee, auf etwa 1900 m über Meer , fand ich in beiden Jahren zwar noch
Jahrgang 123 R. GLATTHAAR. Verbreitung und Ökologie der Kriebelmücken 83
keine Puppen, aber mehrere Altlarven (5. bis 7. Stadium) auf Gras und Wassermoossitzend, Mitte Juni 1977 in zwei gleichartigen Quellbächen auf der Alp Grüm undAlpe Palü unterhalb der Berninapasshöhe (706), ebenfalls auf etwa 1900 m über Meer,drei Larven des 6. Stadiums. Den ergiebi gsten Twinnia führenden Quellbach entdeckteich Mitte Juni 1977 auf etwa 1400 m über Meer. Beim Gasthaus Klausenpass dichtneben der Passstrasse entspringend, führte er Grundwasser, etwa 5 Liter pro Sekunde,und war dicht mit Gras und Wassermoos bewachsen. Auf einer Plastikfolie konnteich neben einigen Altlarven über 250 Puppen und Exuvien ablesen. Be gleitarten warenP. latimucro in etwa gleicher Dichte sowie etwa 80 Larven von E.^cryoplrilum, imOberen gadin trat zu diesen Arten noch E. crenobium. Alle bisheri gen Funde machenes wahrscheinlich, dass T. hydroides reichlich mit Pflanzen bewachsene Quellbächehöherer La gen besiedelt und dass in der Schweiz noch weitere Fundorte zu ent-decken sind.
Die Art, die zu manchen taxonomischen Diskussionen Anlass gegeben hat (RUB-zow, 1959-1964; ROTHFELS, 1956), wird in der vorlie genden Arbeit im Sinne vonKNOZ (1965) und ZWICK (1974) verstanden. Den Erstnachweis für die Schweiz führteL. DAVIES (1957) in seinen alpinen Funden. Wenn auch die Höhenangaben für dieFundorte stark differieren – bei KNOZ (1965) 450 bis 600 m. bei DORIER (1961) 1130bis 2000 m über Meer –, stimmen doch alle Autoren darin überein, dass P. hirtipesam weitesten von allen Prosirnulium-Arten ins Alpenvorland hinab vorstösst. Manfindet sie nicht nur in der montanen Region. sondern gelegentlich auch in Wald-bächen der collinen Stufe, beispielsweise in einem der untersten Seitenbäche der Tössbei der Tössegg (Tö 3). Auch im Tessin fand ich eini ge Larven. Weitere Prosimulium-Funde stammen aus der Westschweiz. Weil es sich aber um nicht identifizierbareJunglarven handelt, habe ich sie nicht ausgewertet. Die in der Thur (Th 10) und inder Töss (Tö 3) gefundenen Einzelgän ger dürften kaum dort aufgewachsen. sonderneher mit der Drift dorthin verfrachtet worden sein.
Die Erstmeldun g lieferte 1927 GALLI-VALERIO aus dem Unterwallis unter demSynonym S. gallü (Eow.kRUS, 1933), die zweite stammt von L. DAVIES (1957). Ichfand die Art in den meisten Alpenbächen und -flüssen. Trotz einer gegenüber anderenArten grösseren Variabilität (KNOZ, 1965) bilden meine Funde ein Taxon, das mitden Befunden von Zwic1 (1974) übereinstimmt. Beim alten Maloja-Hotel (801)
84 Vierteljahrsschrift der Naturforschenden Gesellschaft in Zürich 1973
wurde ich an einem sonnigen Juni-Nachmittag des Jahres 1974 von P. ropes-Weib-chen angeflogen, ohne dass es zu Stichen gekommen wäre. Mitte September fing ichbei der Adula-Hütte, etwa 2000 m über Meer (606c), aus einem Schwarm einigeMännchen.
Diese Art, die nur in Quellbächen höherer La gen gefunden worden ist, ist längereZeit unbekannt geblieben als die anderen Prosimuliuru-Arten. L. DAVIES beschriebsie 1957 aus schweizerischem Material als P. inflaturu; im Jahre 1961 fand sie D. M.
DAVIES zusammen mit T. hydroides im Oberengadin. Wie diese beiden Autoren stellteich an den Fundorten reichlichen Makrophytenbewuchs, relativ geringes Gefälle undgutes Lichtangebot fest.
Obwohl die Art weit verbreitet ist, lie gt noch kein früherer Nachweis aus derSchweiz vor. Ich fand sie in fast allen untersuchten natürlichen Quellbächen der col-linen Stufe. besonders in bewaldeten Gebieten. Sie stei gt auch in die alpine Regionempor. Im Oberengadin besiedelt sie in beachtlicher Dichte den – über 16° C warmen –Auslauf des Lej Nair (804a), in dessen Nähe ich an einem sonnigen Nachmitta g MitteJuni 1974 von E. vernum-Weibchen beflogen wurde, ohne gestochen zu werden. ImMittelland dagegen erwischte ich kein einziges Individuum beim Anflu g oder gar beider Blutmahlzeit, im Gegensatz zu KNOZ (1965) und RUBZOW (1959-1964), welchediese Art als Blutsauger an Tier und Mensch einstufen. Während RUBZOW (1959-1964)fünf Subspezies unterscheidet, die er alle in Russland lokalisiert. ist für Mitteleuropanur mit einem Taxon im Sinne von KNOZ (1965). und ZWICK (1974) zu rechnen.
* Eusimuliurn costatum (FRIEDERICHS 1920)
Fundorte: Li, 214. GK. Gr.530 Larven und Puppen.
So strikte sich die Art auf Waldbäche der collinen Stufe beschränkt, kann dochmit einem geschlossenen Verbreitungs gebiet für Höhenlagen unterhalb 1000 m überMeer gerechnet werden, da sie in den Nachbarländern im selben Habitat an zahl-
Jahrgang 123 R. GLATTHAAR. Verbreitung und Ökologie der Kriebelmücken 85
reichen Orten gefunden worden ist. Die Präima ginalstadien erreichen keine hohenPopulationsdichten, und die Ima gines sind noch nicht beim Blutsaugen beobachtetworden.
Eusimulium crenobium KNOZ 1961
Fundorte: 706, 805.62 Larven und Puppen.
Im Juni 1977 entdeckte ich die Art in den T. hydroides enthaltenden Proben ausdem Oberen gadin. Aus einem kleinen, gut belichteten Quellbach auf der Alp Grüm/Alpe Palü (706) auf der Südseite des Berninapasses, etwa 1900 m über Meer, stammenetwa 50 Larven des 5. bis 7. Stadiums sowie vier Puppen, aus denen zwei Männchenaufgezogen werden konnten, die ZWICK (1977, in litt.) verifizierte. Begleitarten warenneben T. hydroides P. latimucro, E. cr yophilurrr sowie sporadische E. vermin!.
Die Art verhält sich ähnlich wie E. vernum. ausser dass sie in höheren Lagen ihredichtesten Populationen entwickelt. Während sie im Fuldagebiet die häufigste Eusi-muliurn-Art ist (ZwicK, 1974), tritt sie in der Schweiz hinter die beiden zuerst be-sprochenen Eusirnuliurn-Arten zurück. Erst ab etwa 700 m über Meer beginnt siehäufiger zu werden und E. vernum zu ersetzen; im Tessin schon ab etwa 250 m, weildort die montane Region auf diese Höhe hinabreicht. Aufgrund der Dichte der vor-lie genden Nachweise kann im Alpengebiet mit einer geschlossenen Verbreitung ge-rechnet werden. Einzelne Individuen fand ich auch in collinen Waldbächen (Li).
Obschon keine der fündi gen Proben grosse Individuenzahlen enthält, lässt ihregeographische Verteilun g eine gleichmässige Besiedlun g der einschlägigen Biotopeerwarten, zumal da diese Art auch aus den Nachbarländern gemeldet worden ist.
Eusimulium bertrandi (GRENIER und DORIER 1958)
Fundorte: 304, 407.3 Puppen.
Obwohl die Art aus den Westalpen, den Pyrenäen, der Alpensüdseite sowie ausOsteuropa bekannt ist, fehlen noch Nachweise für Zentraleuropa, bis auf zwei Fundeaus der Schweiz. Der eine Fundort liegt bei Trachslau oberhalb Einsiedeln (407), etwa900 m über Meer, wo ich am 25. Mai 1974 ein gut erhaltenes Exuvium mit Kokon inder Alp fand, einem mittel grossen, mässig turbulenten Bergbach, zusammen mitS. rheophilum, P. hirtipes und P. rufipes sowie mit einigen E. vernum und S. variega-
86 Vierteljahrsschrift der Naturforschenden Gesellschaft in Zürich 1978
turn. Zwei weitere Puppen fand ich im Isleterbach bei Isleten (304), der der Alp starkgleicht, zusammen mit E. cryophilurn und O. ornata. Trotz den im Verhältnis zurAnzahl Probestellen in geei gneten Habitaten spärlichen Funden dürften in derSchweiz noch weitere Nachweise möglich sein, da sie auch KNOZ (1965) in schnellen,turbulenten Ber gbächen gefunden hat.
Gruppe Eusirnuliurn angustitarse (LUNDSTROM)
Eusirnuliurn angustitarse (LUNDSTROM 1911)
Fundorte: 409, Gr.3 Larven und Puppen.
Ani 25. Mai 1974 fand ich im Quell gebiet der Biber, in einem Wiesenbächlein inder schwyzerischen Altmatt (409), zwei Puppen sowie im Grendelbach (Gr) eine Alt-larve. KNOZ (1965) und ZWICK (1974) melden aus gleichen Habitaten ergiebigereFunde.
Eusirnuliurn latigoniurr RUBZOW 1956
Fundorte: Gr, Ni, G 4, Hii.Etwa 2500 Larven und Puppen.
Von der Art sind bis heute ausserhalb von Russland erst weni ge Fundorte bekanntgeworden, nämlich aus der Tschechoslowakei (Kwoz, 1965), dem Apennin (Rivo-SECCHI, 1967) sowie je ein Fund aus England (L. DAVIES, 1966) und aus dem Fulda-gebiet (ZwicK, 1974). Aufgrund der vorliegenden Aufsammlungen gehört sie im Kan-ton Zürich zu den häufi gen Eusirnuliurn-Arten. Wie die genannten Autoren überein-stimmend berichten, leben die Präimaginalstadien in kleinen, nur mässi g rasch flies-senden und daher im Sommer warm werdenden, stark mit Pflanzen zugewachsenenBächen und Entwässerungsgräben. Im Kanton Zürich passt die Beschreibung auf denGrendelbach (Gr) und den Niderbach (Ni), welche die dichtesten Populationen be-herbergen. Sporadisch fand ich E. latigoniwn im Hüttwilerseebach (Hü) sowie in derGlatt bei Niederglatt (G 4); die Art besiedelt somit vor allem Ausläufe kleinerer Seenoder von Riedgebieten. Ausserhalb des Kantons Zürich und des angrenzenden Thur-gaus gibt es noch keine Funde.
Die E. aureum-Gruppe umfasst einige nahe verwandte Arten, die nur anhand derGenitalien identifiziert werden können. Vertreter dieser Gruppe fand ich als Larvenoder Puppen an verschiedenen Orten der AlpeHnord- wie auch der Südseite. Puppen-funde, aus denen Ima ginalzuchten gelangen, lie gen aus dem Niderbach (Ni), aus demHüttwilerseebach (HU) sowie aus dem Auslauf des Lej Nair (804a) vor. Die Genitalien
Jahrgang 123 R. GLATTHAAR. Verbreitung und Ökologie der Kriebelmücken 87
stimmen mit den Abbildun gen überein, die RUBZOW (1959-1964) von E. securiformeRuszow 1956 = E. angustipes (EDWARDS 1915) (Zwick, 1974) herausgab. Am nörd-lichen Stadtrand von Winterthur fing ich am Abend des 16. August 1973 mit einemSchwarm von etwa 1000 W. lineata-Weibchen auch eines von E. angustipes, in einerEntfernung von über 10 km vom nächsten nach gewiesenen Brutplatz, in einem jauche-haltigen Wasser graben bei Thalheim an der Thur, etwa zwei km westlich von Th 10.
Tribus Wilhelmiini RUBZOy 1974
Genus Wilhelmia ENDERLEIN 1921
Wilhelmia equina (LINNAEUS 1747)
Hauptfundorte: G 5, Tö 2, Tö 3, ZB, Fl, 207, 211, 217, G 2 bis G 4, Rh, 120b, 231,509, Th 10.Etwa 8300 Larven und Puppen.
W. equina ist in allen Teilen des europäischen Flachlandes häufig und wird seitlanger Zeit als Blutsau ger an Tier und Mensch gefürchtet. In der Westschweiz ist sie,auch wenn die ungleiche Probestellendichte berücksichtigt wird, seltener als in derOstschweiz, insbesondere im Kanton Zürich, wo mehrere ergiebige Brutplätze bekanntgeworden sind; es handelt sich durchwe gs um eutrophe, wenig turbulente, grössereBäche oder Flüsse. KUHN (1952) meldet einen nicht verifizierbaren Fund aus dembedeutend kleineren Stadtzürcher Wehrenbach, der zum grossen Teil Waldgebietdurchfliesst. Im Sommer 1973 fing ich einige Dutzend Weibchen beim Anflug ansVieh.
x Wilhebnia lineata (MEIGEN 1804)
Hauptfundorte: 117, Rh, Th 10, 120a, G 2 bis G 4, 119a, 121, 235, 415, 203, 217,229, 509, Th 9.Über 28000 Larven und Puppen.
Die Art fand ich nicht, wie RUBZOW (1959-1964), in raschen, kühlen und klarenBächen der Submontanstufe, sondern in lan gsameren, eher trüben Bächen und See-ausflüssen des Flachlandes, die sich im Sommer auf über 20' C erwärmen können,wie RUBZOW für das Synonym W. salopiensis angibt. Die Nachweise von W. lineatakonzentrieren sich noch stärker auf die Nordostschweiz als jene von W. equina. Ob-wohl sie in der Thur (Th 10) und im Hochrhein (Rh, 117, 119a, 120a, 121) jedenSommer grosse Populationsdichten erreicht und ihre Frühsommer generation zu Zehn-tausenden übers Weidevieh herfällt, fehlte sie bis jetzt in der schweizerischen Faunen-liste. Ende Juni 1973 fielen in Thalheim an der Thur vier Rinder ihren Stichen zumOpfer (ECKERT, unpubliziert). Der Mensch wird ebenfalls umschwärmt; Blutsaugenkonnte ich aber nicht beobachten. Nachdem auch aus Rheinfelden und MuttenzKlagen über Kriebelmücken-Anflüge laut geworden sind, ist anzunehmen, dass derHochrhein bis Basel besiedelt ist, soweit nicht Stauwehre den Fluss hemmen. Unter-halb von Basel scheint der Rhein wohl infolge der starken zivilisatorischen Belastungfrei von Simuliiden zu sein (GRUNEWALD, 1972).
88 Vierteljahrsschrift der Naturforschenden Gesellschaft in Zürich 1978
Tribus Simuliini GRENIER und RAGEAU 1960
Genus Boophthora ENDERLEIN 1921
Boophthora erythrocephala (DEGEER 1776)
Hauptfundorte: 323, Hü, 230, G 1, 204, 612, 229, G 2 bis G 4, Rh, 415, 416, 622,Ni, 203, 420 a.Über 35000 Larven und Puppen.
Weil B. erythrocephala im Aller-Leine-Gebiet schon seit Jahrzehnten als Blutsaugerberüchtigt ist, wird ihre Biolo gie mit Unterbrüchen seit 1920 (WILHELMI) extensiv, seit1967 (RUEHM et al.) intensiv untersucht. Sieht man von der nicht verifizierbaren An-gabe von KUHN (1952) für den Wehrenbach ab, so verdanken wir den Erstnachweisder Art für die Schweiz ECKERT et al. (1969), die durch die Todesfälle von Rindernbei Schwerzenbach auf den 1968 offenbar besonders dicht besiedelten Brutplatz G 1aufmerksam wurden. Auch 1973 beteiligte sie sich an den Attacken auf das Vieh,kann aber wegen ihrer gegenüber W. lineata zehnfach geringeren Individuenzahl nichtin erster Linie für die Todesfälle bei Thalheim verantwortlich gemacht werden. Da-gegen kam es bei Expositionsversuchen zu Anflug und Stichen am Menschen, wobeiB. erythrocephala die Unterschenkel sowohl der stehenden wie auch der sitzenden, nurmit Turnhosen bekleideten Versuchsperson am häufi gsten an griff. Auch Unterarmeund Gesicht waren Ziele ihrer Anflüge. Innert 15 Minuten konnte ich 50 Weibchenabsammeln, die zum Stich angesetzt hatten. Alle gehörten zu B. erythrocephala, keineszur gleichzeitig in der Überzahl anflie genden Gattung Wilhelrnia. Wichti gste Brut-stätten von B. erythrocephala im nördlichen Kanton Zürich und angrenzenden Thur-gau sind der Hochrhein (120 a) und der Hüttwilerseeauslauf (Hü), wo die Mücken.ihre Larven und Puppen am reichlichen Schilfbewuchs viel Eiabla ge- und Anheftungs-substrat vorfinden, das ihnen Besiedlungsdichten bis 5 ermö glicht. Weitere Fundortebefinden sich in der Limmat, in der Aare sowie vor allem in Seeausflüssen, unterdenen die Lorze bei Cham (323) besonders dicht besiedelt war. Auf der Alpensüdseitefand ich die Art in der Tresa (622) sowie in einem kleinen, trägen und stark ver-krauteten Lan genseezufluss, der links neben dem Ticino bei Maltadino in den Zan gen-see mündet (612). Höchst gelegener Fundort ist der Auslauf des Hüttenersees (420)auf etwa 650 m über Meer.
Odagrrria ornata ist die häufi gste Kriebelmückenart der Schweiz. Im Mittelland undJura fehlt sie in fast keinem Bach oder Fluss und entwickelt ihre dichtesten Popula-tionen in kleineren Wiesenbächen, einem der häufi gsten Gewässertypen der Schweiz.Mehr als ein Drittel aller gesammelten Kriebelmücken gehört zu dieser Art. Diehöchsten Fundorte liegen im Oberengadin: im Auslauf des Lej Nair (804 a) und im
Jahrgang 123 R. GLATTHAAR. Verbreitung und Ökologie der Kriebelmücken 89
Inn bei Samedan (806), der den St. Moritzersee entwässert. Diese Gewässer sind keinetypischen Ber gbach-Habitate, sondern gleichen eher den Flachland gewässern (hoheSommertemperaturen, geringe Fliess geschwindigkeit und Turbulenz, starker Pflanzen-bewuchs). Auf der Alpensüdseite ist sie in der Magadinoebene (Seitenbäche desTicino) und im Mendrisiotto häufi g. O. ornata wird auch in der Schweiz ab und zulästig. So berichtet GALLI-VALERIO bereits 1927, bei der Erstmeldung für die Schweiz,dass Weibchen der Art in Lausanne Menschen beflo gen haben. Meine Anflugbeob-achtungen umfassen einige Weibchen, die im Oktober 1973 ums Vieh schwärmten,sowie solche, die Mitte März 1974, an den ersten warmen Frühlingstagen des Jahres,mich anflo gen und zu stechen versuchten. Weil O. ornata als Artkomplex angesprochenworden ist (Ruszow, 1959-1964; RUEHM, 1967), führte ich an mitteleuropäischemMaterial eine morphometrische Analyse durch, die indessen keine Anhaltspunkte füreine taxonomische Heterogenität ergab (GLATTHAAR, in Vorbereitung).
Diese nächste Verwandte von O. ornata kann nur auf dem Puppenstadium anhandder zahlreichen, spitzen Thorakaltuberkel zweifelsfrei von der Schwesterart unter-schieden werden. ZwicK (1974) wies sie in der Gryonne bei Bex (506) nach, ein Fund,den ich 1974 bestätigen konnte. Weitere Fundmeldungen lie gen aus allen Nachbar-ländern vor; alle Autoren haben sie in Quell- und Waldbächen gefunden. Bei Einzel-funden in grösseren Flüssen (Reuss bei Mellingen [320] und Töss [To 2]) dürfte essich um Individuen handeln, die aus den genannten Habitaten verdriftet worden sind.Auch in den grösseren Alpenflüssen ist sie selten, während kleinere Voralpen- undJurabäche die grössten Populationen beherbergen. Gerade am Jurasüdfuss scheint siedie häufi gste Art zu sein. In einem kleinen, schnell fliessenden Bächlein beim Restau-rant Glutzenberg oberhalb von Günsberg (214) ersetzt sie O. ornata in ähnlichenPopulationsdichten. Dort fand ich an Grasblättern und Zweigen auch Eiklumpen,die ich O. spinosa zuschreibe. In ähnlicher Dichte besiedelt sie einen Zufluss desFätschbaches auf dem Urnerboden (401), wo ich im Juni 1977 etwa 700 Larven undPuppen sammelte. Puppen fand ich von April bis Oktober, ohne ihre Generationen-zahl angeben zu können. Auch über das Verhalten der Ima gines liegen noch keineBeobachtungen vor.
Genus Tetisiinuliurn RuBZOw 1963
*Tetisirttulium bezzii (CORTI 1916)
Fundorte: 503 b. 210, 209.27 Larven und Puppen.
Obwohl die Art als zirkummediterraner Endemit gilt (Ruszow, 1967; ZWICK,
1974), konnte ich sie für die Schweiz und zu gleich für die «Re gion 9» der «Limno-fauna Europaea» im August 1973 erstmals nachweisen, nämlich in der Gerine beiMarly-le-Grand im Üchtland (210), einem mittelgrossen Voralpenfluss mit relativ
90 Vierteljahrsschrift der Naturforschenden Gesellschaft in Zürich 1978
seichtem, turbulentem Wasser, der die Sohle eines kleinen Tobels einnimmt. Von derProbe, die nur wenige Individuen umfasste, gelang mir aus einer Puppe die Aufzuchteines Männchens, dessen Identität ZWICK (in litt.) bestätigte. Im Rahmen der Quer-schnittsuntersuchung machte ich zwei weitere Funde, den einen in der Saane bei LaTine (209), einem Ort, der dem genannten stark gleicht, einige Puppen und Exuvien,den anderen im Wallis in der Bor gne bei Bramois (503 b), mit 17 Larven und Puppenden ergiebigsten. Vermutlich markieren diese Funde die Nordost grenze des Verbrei-tungsgebietes.
S. variegatum lebt in grösseren Bächen und Flüssen der Submontanstufe, besiedeltpflanzliches, aber auch anorganisches Substrat und hält sich im all gemeinen unter1000 m über Meer. Die Art wird für gleichartige Habitate aus Mittel-. West- undSüdeuropa sowie aus Grossbritannien gemeldet, nicht aber aus Nordeuropa. Den fürdie Schweiz ersten und zu gleich ergiebigsten Fund machte ich am 1. Juli 1973 in derSihl bei Sihlbrugg (412), wo Larven und Puppen in grosser Dichte siedelten undO. ornata fast ganz fehlte. Puppen fand ich von Mai bis Oktober; KNOZ (1965) undZWICK (1974) geben zwei Generationen an. Im Oktober fand ich bei Sihlbru gg Jung-larven, die ich dieser Art zurechne, und nehme an, dass sie auf dem Larvenstadiumüberwintert. Im Frühjahr 1974 fing ich im unteren Tösstal auf einer mit Rindernbestossenen Weide einige Weibchen, so dass die Art auch in der Schweiz Blut saugendürfte.
Gruppe Simulium monticola FRIEDRICHS
Simulium monticola FRIEDRICHS 1920
Hauptfundorte: 129, 112, 406b, Th 1.255 Larven und Puppen.
Trotz reichlichen Funden der Art in den Nachbarländern – ZWICK (1974) beob-achtete so gar ihre Eiabla ge – erbeutete ich selten mehr als vereinzelte Individuen proSammelstelle. Als er giebigste erwiesen sich die Proben aus dem Scheltenbach beiSchelten (129) aus dem Solothurner Jura, sowie aus Wiesenbächlein bei Klosters undauf der Alp Schlappin oberhalb Klosters (112) im Herbst 1973. Die Nachweise stüt-zen sich auf Puppenfunde, da die Larven nicht mit genügender Sicherheit von jenender nahe verwandten Arten S. rheophilum, S. maximum und S. variegatum zu unter-scheiden sind. Anders als KNOZ (1965) und ZWICK (1974) fand ich S. monticola amehesten in kleineren Wiesenbächen und Quellrinnsalen der Montanstufe; sie scheint
Jahrgang 123 R. GLATTHAAR. Verbreitung und Ökologie der Kriebelmücken 91
auch weniger weit ins Mittelland hinab vorzustossen als S. variegation und S. rheo-
Im Unterschied zu ZWICK (1974) fand ich die Präima ginalstadien der Art, wieKNoz (1965), in grösseren, wasserreichen und turbulenten Bergbächen, wenn auchmeist nur in geringer Dichte. Nur in der Alp oberhalb Einsiedeln, bei Trachslau (407),erreichte sie hohe Dichtegrade, wo ich am 25. Mai 1974 eine Probe nahm. Die meistenPuppen sassen auf faust- bis kopfgrossen Geröllen. Höhenlage und Bachtypus gleichendem Habitat von S. variegaturn.
Sirnulium maxirnum (KNoz 1961)
Hauptfundorte: 303, 301, 601b, 801, 602b.214 Larven und Puppen.
Von der Art liegen auch aus den Nachbarländern erst wenige Fundmeldungen vor.Wie KNoz (1965) fand ich sie in grösseren, raschen und turbulenten Gebirgsbächenund -flüssen, in Gesellschaft mit P. rufipes uHd S. rheophilum, sowie auch in kleineren
Seitenbächen, zusammen mit S. rnonticola. Ergiebi gste Fundorte waren der Schächen-bach (303), wo ich Anfang Juli 1974 auf Ästen und Steinen zahlreiche Puppen fand,sowie das Gotthardgebiet (301. 601 b).
Gruppe Sirnulium tuberosum (LUNDSTROM)
* Sirnulium tuberosurn (LuNnsTROM 1911)
Fundorte: 601 a, 410, 411, 322.340 Larven und Puppen.
Die Art wird aus Mittel- und Nordeuropa als häufiger Bewohner grösserer, relativlan gsamer, tiefer und turbulenzarmer Mittelgebirgsbäche gemeldet (KNoz, 1965;ZWICK, 1974), ferner aus den Pyrenäen (GRENZER, 1953) sowie aus dem Apennin
(RlvosEccxt und LIPP.ARONI. 1965), welche Funde etwa die südliche Verbreitungs-
grenze markieren dürften. Die Erstfunde für die Schweiz beschränken sich auf vierOrte, von denen nur einer ergiebi g war. Es handelt sich um den Ausfluss aus dem
Lar go Cada gno, einem relativ warmen und rasch fliessenden kleineren Zufluss zumRitomsee oberhalb von Airolo (601 a), die mit 1900 m über Meer mit Abstand höchst-gelegene Fundstelle. Vereinzelte Larven und Puppen fand ich in der Lorze bei Baar(322), wo S. variegation dominierte, ferner in der Alp bei Biberbrugg, vor der Ein-mündun g in die Biber (410), auf Steinen, und schliesslich in einem Wiesenbach inEgg bei Einsiedeln (411) eini ge Altlarven. Diese Habitate der kleinsten schweizerischenSimuliidenart entsprechen den ausländischen weitgehend.
92 Vierteljahrsschrift der Naturforschenden Gesellschaft in Zürich 1978
Die zahlreichen Fundmeldun gen für die Art betreffen fast ausschliesslich Teich-und kleinere Seeausflüsse, wo sie sich auf die vom Wasser überströmten Pflanzen,Dammkronen, Steine sowie die Gegenstrudel der Kolke konzentriert. An solchenOrten entwickelt sie maximale Populationsdichten. Larven und Puppen sitzen oftmehrschichti g übereinander und bilden Klumpen, erweisen sich dennoch als driftfest.Ergiebi gste Fundorte sind der Lützelseeauslauf (Lü, Dammkrone), die Schleuse desHüttwilerseeauslaufes (420b), der Lucelle-Teichausfluss (127, Abflussschacht undKaskade), der Bollement-Teichausfluss (128, ein mit Fontinalis bewachsener Kanal)sowie, als höchstgelegener Fundort, der Hahnenseeauslauf auf 2150 m über Meer(804b), an Steinen oberhalb der ersten Stromschnellen. In wenig tnrbulenten See-ausflüssen tieferer Lagen kommt S. argyreatum nur sporadisch vor, am zahlreichstenim Grendelbach (Gr). ZWICK und RUEHM (1972) berichten über Blutsausen der Som-mergeneration.
Obwohl die Art zu den ältesten bekannten Simuliiden gehört und in Mitteleuropanicht selten ist, liegt aus der Schweiz noch keine Fundmeldun g vor. Ähnlich wie beiden beiden Wilhelmia-Arten fand ich die grössten und dichtesten Populationen in derNordostschweiz, in relativ seichten, mässi g raschen, turbulenten und im Sommer sicherwärmenden Flüssen und grösseren Bächen des Alpenvorlandes. HöchstgelegenerFundort ist der Silsersee-Ausfluss (803 b). Die Weibchen werden in manchen Gegen-den Europas als Blutsauger gefürchtet. So identifizierte ich neben B. erytlrroceplralaund W. lineata auch einige Individuen als S. reptans in den Kriebelmückenproben,die das Institut für Parasitolo gie der Universität Zürich von den Ende Juni 1973 beiThalheim verendeten Rindern genommen hatte. Dennoch fing ich aus den Schwärmenmit dem Kescher nie mehr als vereinzelte Exemplare – am meisten am 28. April 1975 –und wurde nie von S. reptans gestochen.
Einzelfunde sind aus allen benachbarten Alpengebieten gemeldet worden. Aus derSchweiz liegt nun ein reichhaltiges Material vor. Nicht nur konnte ich die Art für die
Jahrgang 123 R. GLATTHAAR. Verbreitung und Ökologie der Kriebehnücken 93
meisten Ber gbäche und Bergflüsse nachweisen, sondern stiess auch auf Populationenhoher Dichte grade in der En gelberger Aa bei der Titlisbahnstation in Engelber g (309),im Ticino bei Giornico (603), im Brenno bei Acquarossa (606a), im Oberlauf derThur (Th 1 bis 6), in der Kleinen Emme (314, 316) sowie auch in der Mera bei Piuro(703). Trotz des reichlichen Vorkommens der Präimaginalstadien konnte ich keineImagines beim Schwärmen oder beim Anflug beobachten.
Slrrtuhurn degrangei DORIER und GRENIER 1959
Fundorte: Th 4 bis 10, sowie grössere Zuflüsse.Über 400 Larven und Puppen.
Von der Art, welche ähnliche ökologische Ansprüche zu stellen scheint wie S. ar-genteostriatum, liegen erst wenige Fundmeldun gen vor: Französische Alpen (DORIER
und GRENIER, 1959; DORIER, 1961), Italien (RIvosEccHi, 1963, 1967), Jugoslawien(ZIVKOVlrcx, 1961) sowie Tschechoslowakei (KNOZ, 1963). In der Schweiz fand ichsie nur in der Thur und in den unteren Partien ihrer grösseren Nebenflüsse im mitt-leren Toggenburg, wo die Larven und Puppen in grosser Zahl auf Steinen und ver-holzten Zweigen sassen. Die meisten Puppen fand ich im Frühjahr, die letzten EndeJuli, im Unterlauf bereits Ende Mai. Der höchstgelegene Fundort ist Th 4, 740 m überMeer. Da S. degrangei auf dem Larvenstadium nicht von S. argenteostriatum unter-schieden werden kann, die an allen Fundstellen ebenfalls vorkommt, konnte ich ihreEiablageplätze nicht lokalisieren. KNOZ (1965) meldet einen etwa 1000 m über Meergelegenen Fundort.
4. Phänologie der Simuliidae
Die Lebenszyklen von 24 mitteleuropäischen Simuliidenarten wurden von ZWICK
(1974) anhand von Material aus dem Fuldagebiet dargestellt. Deshalb kann ich michim fol genden auf ergänzende An gaben beschränken.
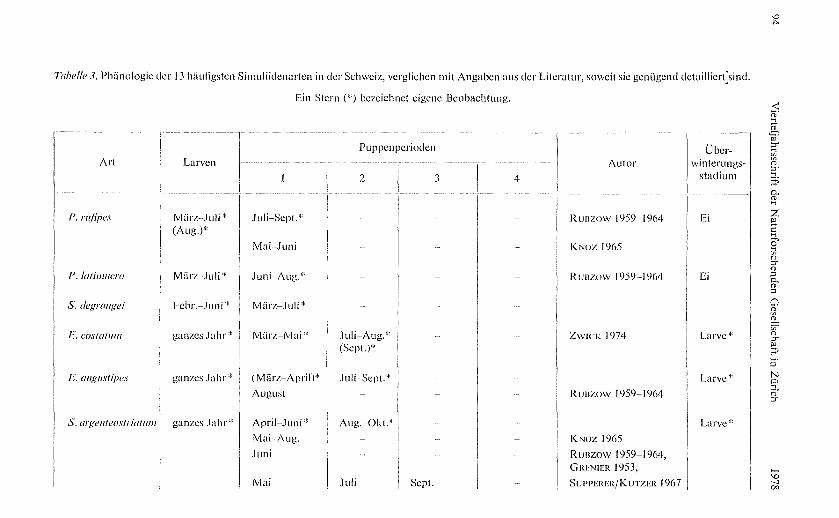
Tabelle 3 gibt Auskunft über das zeitliche Auftreten von Larven und Puppen der13 am häufi gsten gefundenen Arten anhand des 1973 und 1974 gefundenen Materials,dargestellt über ein Kalenderjahr hinwe g und geordnet nach zunehmender AnzahlGenerationen.
Die Prosimulium-Arten durchlaufen auch nach meinen Beobachtungen nur eineGeneration im Jahr. Bei P. rufipes zieht sich die Puppenzeit am län gsten hin. P. hir-tipes, eine Art, die auch in tieferen Lagen vorkommt, verpuppt sich schon im Früh-jahr. Da im Herbst keine Prosirnulium-Larven gefunden worden sind, ist anzunehmen,dass die Gattun g im Eistadium überwintert.
Die Eusimulium-Arten unterliegen von Zyklus zu Zyklus den grössten Bestandes-schwankungen; ihre Populationsdichten sind fast durchwegs gering, und die K lima-faktoren wie auch die hydrologischen Bedingun gen dürften einen stärkeren Einflussausüben. Während E. vernum im Frühjahr die ergiebigsten Funde lieferte, tendiertendie übrigen Arten der Gattung zu Dichtemaxima im Sommer.
Beide hVilhelmia-Arten waren im Frühjahr in relativ geringer Populationsdichte zu
Tabelle 3. Phünologie der 13 häufigsten Simuliidenarten in der Schweiz, verglichen mit Angaben aus der Literatur, soweit sie genügend detailliert'sind.
Ein Stern ("') bezeichnet eigene Beobachtung.
Art Larven
Puppenperioden
2 3 4Autor
Über-winterungs-
stadiunm
P. rufipes März-Juli'" Juli-Sept." RUnzow 1959-1964 Ei(Aug.)*
Nlai---Juni KNOZ 1965
Y. latimucra Miirz--Juli" Jnni--Aug.''' RutIZOW 1959-1964 Ei
S. degrangei (Febr.-Juni '` März-Juli"
L. casurtum ganzes Ja li r'* -Mai* Juli-Aug.'s Zwici. 1974 Larve*(Sept.)*
Miirz-April "` Juni"' August * Okt.(-Nov.)'" Larve*
April-Mai August September ZWICK 1974,0 RUNEWALD 1972
96 Vierteljahrsschrift der Naturforschenden Gesellschaft in Zürich 1978
finden, während besonders die Frühsommergeneration von W. lineata so zahlreicheIndividuen hervorbrin gen kann, dass deren Stechaktivität zu Viehschäden führt.
O. ornata und B. erythrocephala brachten 1973 vier Generationen hervor, im fol-genden Jahr – wohl wegen des verfrühten Wintereinbruches – an höher gelegenenBrutplätzen nur drei. Im Spätjahr kam es bei den Puppenperioden zu Überlappungen,besonders bei O. ornata, während jene von B. erythrocephala besser synchronisiertschienen, auch besser als bei den B. erythrocephala-Populationen im Aller-Leine-Ge-biet (RUEHM, 1971). Während RUEHN1 (1971) und ECKERT et al. (1969) die Frühjahres-generation als stärkste und fürs Vieh gefährlichste nach gewiesen haben, verlegte sich1973 und 1974 das Schwer gewicht auf die zweite oder dritte Generation, in der Glattbei Schwerzenbach vermutlich deshalb, weil die Schilfbestände im Frühjahr ge-schnitten wurden.
In den relativ warmen Seeausläufen des Oberen gadins passen sich Arten, die auchim Tiefland vorkommen. dem stren geren Klimare gime und dem kürzeren Sommeran. Mitte Juli und Mitte Oktober 1976 durch geführte Exkursionen erbrachten imSommer von O. ornata und S. argyreatum Altlarven, Puppen, Exuvien, Gelege undJunglarven, im Oktober von O. ornata dieselben Stadien, von S. argyreatum Alt-larven, Puppen. Exuvien und an Steinen grosse Men gen von Eiern. Somit durchliefenbeide Arten zwei Zyklen mit gleichen Verpuppungsterminen. Die Eusirnulium-Artendürften ihrerseits nur eine Generation hervor gebracht haben, da ich im Juli vonE. vernum und E. angustipes Altlarven und Puppen fand, im Oktober von E. vernumnur einige Jun glarven, von E. angustipes nur eine Puppe. Diese Art dürfte auf demEistadium überwintert haben, nachdem Anfan g November die ersten ergiebigenSchneefälle den Winter gebracht haben.
5. AssoziationeH zwischen Simuliidenarten
Nachdem in den meisten Proben mehr als eine Kriebelmückenart enthalten ist– die Anzahl Proben mit k = 1, 2, ... S Arten ist annähernd poissonverteilt –‚ stelltsich die Fra ge, welche Kriebelmückenarten häufiger miteinander in derselben Probegefunden worden sind, als es einem zufälli gen Zusammentreffen entspräche. SolcheFälle werden als Assoziationen bezeichnet.
Tabelle 4 gibt die Anzahl Funde der 17 Arten mit mehr als 30 Nachweisen aus derLangzeit- und aus der Querschnittsuntersuchung. Mit Hilfe der Chiquadrat-Kontin-genztafel lässt sich für jeden kombinatorisch sich er gebenden Artpaarwert die Hypo-these testen, dass beide Arten unabhängig voneinander vorkommen (DicE, 1945). Alsassoziiert bezeichnen wir Artpaare. deren Chiquadratwert grösser ist als 6,41 (ent-spricht p<0,01). Es ist zu beachten, dass nicht alle Daten unabhängig voneinandersind, weil an vielen Probestellen mehr als eine Probe genommen worden ist, wennauch in einem Zeitabstand von sechs Wochen oder län ger, und weil die mehr als zweiArten enthaltenden Proben nach Artpaaren aufgeschlüsselt worden sind, woraus sicheini ge Überinterpretationen ergeben. Zudem ist das Er gebnis auch der Grösse undAnzahl der Stichproben unterworfen, und bei stark verschiedenen Fundhäufigkeiten
Jahrgang 123 R. GLATTHAAR. Verbreitung und Ökolo g ie der Kriebelmücken 97
Tabelle 4. Anzahl Funde der 17 häufigsten Arten in den insgesamt 774 fündigen Proben
(beispielsweise O. ornata gegenüber W. equina) besteht die Assoziation nur einseitig,bei der selteneren Art (D1cE, 1945, 1948; MÜHLENBERG, 1976).
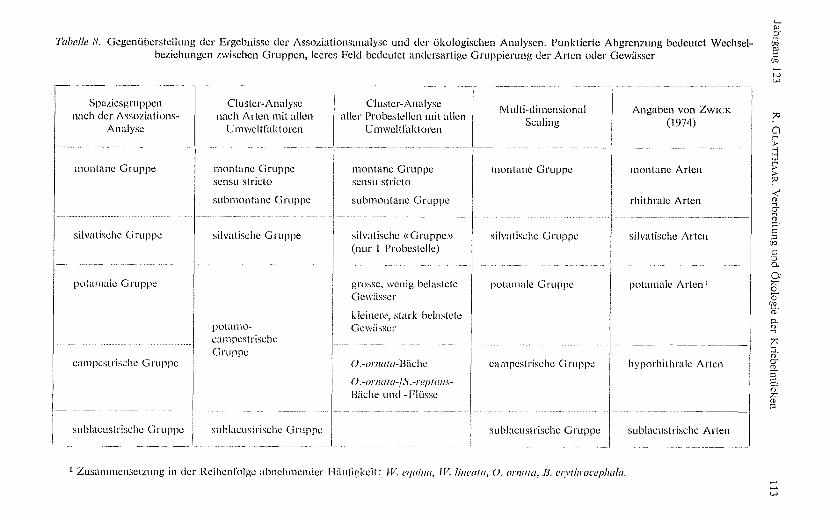
Aufgrund dieser Berechnun gen lassen sich fünf Gruppen abgrenzen, die mit demsubjektiven, während der Sammeltätigkeit im Felde gewonnenen Eindruck überein-stimmen. Innerhalb einer Gruppe ist nicht jedes Mitglied mit jedem assoziiert. Sosind P. rufipes und S. argenteostriatum indirekt, über E. carthusiense, miteinander ver-bunden, während die Frequenz ihres direkten Zusammentreffens zufällig erscheint.Da sich aus den Gruppen kein direktes, kausales Verständnis der Verteilung undAbundanz der Arten ergibt, sind sie nur als erster Versuch einer Gruppierung zu ver-stehen, die für die fauvistisch-ökolo gische Forschung einerseits, für die angewandteveterinärmedizinische Arbeit anderseits von Nutzen sein kann.
Abbildung 3 visualisiert die Assoziationen und Gruppen, die ich benenne mit
I. montane Gruppe, Bergbachbewohner: P. rufipes, E. carthusiense, E. cryo-philurn, S. variegatum, S. rheophilurn, S. argenteostriatum;
II. silvatische Gruppe, Waldbachbewohner: E. costatum, E. vernum, O. spi-nosa sowie, mit Beziehung zur Montan gruppe, E. cryouphilum;
III. potamale Gruppe, Bewohner g rösserer Flachlandflüsse: W. equina,W. lineata, B. erythrocephala;
IV. campestrische Gruppe, Bewohner von Feld- und Wiesenbächen:O. ornata, S. reptans sowie, mit Beziehungen zu III, W. equina;
V. sublacustrische Gruppe, Seeausflussbewohner: E. latigoniurrr, E. an-
gust ipes, S. argyreaturn.
Neben diesen Hauptbeziehun gen lassen sich auch eini ge schwache Assoziationenerkennen. Der wichtigste Überschneidungsraum ist das Voralpengebiet, wo P. rufipes
98 Vierteljahrsschrift der Naturforschenden Gesellschaft in Zürich
1978
Abb. 3. Anordnung der 17 häufi gsten Simuliidenarten anhand des «Muttidimensional Scaling>, mitden Populationsdichten je Art in den 350 ergiebigsten Proben. Die Skalen auf der Ordinate und aufder Abszisse geben ein relatives Ähnlichkeitsmass. Ferner sind eingetragen die Assoziationen zwischenden Arten. Die dicken Verbindun gslinien bedeuten hochsi gnifikante (p,0,001), die dünnen Liniensignifikante (0,01 >p>0,001) Assoziation. Mit gestrichelten Linien sind die subjektiv vorgeschlagenen
Gruppen umgrenzt.
und E. vernum einerseits, S. argenteostriatum und S. reptans anderseits häufig zusam-men gefunden worden sind, gefolgt vom Jura, wo sich die beiden Odagmia-Artenbege gnen. Wo Seeausflüsse in rhithrale Wasserläufe übergehen, treffen O. ornata,E. latigonium und E. angustipes aufeinander, wo sich ein Potamal anschliesst, B. ery-throcephala und E. angustipes. Die nur O. ornata enthaltenden Proben stammen ausden zahlreichen Wiesenbächen, zum Beispiel der Kempt, dem Altbach, dem Sennhof-bach. In breiteren Bächen wie etwa der Töss unterhalb von Winterthur tritt S. rep-tans hinzu.
Auch eini ge sublacustrische Habitate sind nur von einer Art besiedelt. Im Mittel-land fand ich in langsamen und turbulenzarmen Seeausflüssen B. erythrocephala, inraschen und turbulenten dage gen S. argyreatum, im Oberen gadin in den langsamenAusflüssen vornehmlich O. ornata und E. angustipes, in den raschen S. reptans, E. ver-nunr und O. ornata. S. argyreatum kam in beiden Biotopen vor.
Berücksichtigt man bei den 350 in bezug auf Arten- und Individuenzahlen ergiebig-
Jahrgang 123 R. GLATTHAAR. Verbreitung und Ökolo gie der Kriebelmücken 99
sten Simuliidenproben die geschätzte Besiedlun gsdichte je Art, so ergeben sich einigeAbweichun gen zur Assoziationsanalyse. Als Gruppierungsverfahren bietet sich dieCluster-Analyse an. Als Operational Taxonomic Units (OTUs) dienen die 17 Arten,als «Merkmale» die Populationsdichte-Werte von 0 bis 5. Die Werte wurden je Merk-mal auf die Standardabweichun g s = 1 standardisiert und die taxonomische – hierökolo gische – Distanz d; k: zwischen zwei OTUs berechnet nach
m1 `^
(Xlj —Xik)?m1
wobei Xi; der standardisierte Wert des Merkmals i für OTU j, Xik für OTU k undm die Anzahl der Merkmale ist. Aufgrund der daraus resultierenden Distanz-Matrixlassen sich die Taxa nach ihrer relativen Ähnlichkeit gruppieren. Von dieser Matrixausgehend, liefert das Verfahren «Multidimensional Scalin g » ein zweidimensionalesKoordinatensystem, dessen Skalen ein relatives Mass für die Ähnlichkeit in allenuntersuchten Merkmalen darstellen. Abbildung 3 zei gt, dass sich die meisten Simu-liidenarten um das Zentrum scharen; immerhin lassen sich die fünf Gruppen aus derChiquadrat-Analyse teilweise wiedererkennen. Im Zentrum finden wir, erwartungs-gemäss nahe beisammen, die montanen und die silvatischen Arten; zur Linken diesublacustrischen und zur Rechten die potamalen Arten, während die beiden cam-pestrischen an den oberen Rand zu lie gen kommen; O. ornata überschreitet die Skalaum mehr als eine Einheit. Diese Abweichung dürfte auf die stark variierenden Popu-lationsdichten und ein euryökes Verhalten der beiden campestrischen Arten zurück-zuführen sein, was ihnen ein gemeinsames Vorkommen mit fast allen anderen Artenermö glicht (Abb. 3). Auf der anderen Seite weisen auch ausgesprochene Biotop-spezialisten wie S. argyreatuin, E. latigouium sowie B. erythrocephala grössere Ab-weichungen auf, vor allem in der horizontalen Komponente. Demgegenüber fällt diezentrumsnahe La ge von W. equiva auf, ist doch die Art mit ihren Nachbarn S. varie-gatum, S. rheophilum und S. argenteostriatum insgesamt nur fünfmal zusammen gefun-den worden. Diese Analyse ist demnach empfindlicher auf Abweichun gen vorn Mittel-wert als die Chiquadrat-Analyse, welche nur Vorkommen oder Nichtvorkommenberücksichtigt. S. reptans und O. ornata scheint sie mir im Hinblick auf die bespro-chenen Assoziationen zu stark von den anderen im Mittelland vorkommenden Artenabzusondern. Neben phänologischen Gegebenheiten und der Auswahl der Probendürfte wie bei der Assoziationsanalyse auch das Verhältnis fündi ger zu nichtfündigerProben je Art eine Rolle spielen. Somit darf auch die Gruppierung nach dem Multi-dimensional Scaling nur aus kritischer Distanz betrachtet werden.
Untersucht man einen grösseren Fluss wie die Thur von der Quelle bis zur Mün-dung auf seine Simuliidenfauna, so zeigt sich ein zur vorgeschla genen Gruppierungpassendes Artenspektrum. Meine zehn Probestellen entsprechen in ihren Abständenetwa gleich grossen makroskopischen Veränderungen in der Gewässermorphologie.Ende September 1973, Anfan g Juni 1974 und Ende September 1974 nahm ich an jederStelle mindestens eine Probe. Tabelle 5 gibt die Anzahl der gefundenen präimaginalenIndividuen. Sie zei gt, dass einige Arten in bestimmten Flussabschnitten dominieren,das heisst mindestens 50% der IHdividuen stellen:
1/2
d, 1. =
100 Vierteljahrsschrift der Naturforschenden Gesellschaft in Zürich 1978
Th 1 bis Th 2: P. rufipes, vergesellschaftet mit S. argenteostriatum, E. cryophilurn,E. car tlrusiense, S. monticola, P. latimucro.
Th 2 bis Th 5: S. argenteostriatum, vergesellschaftet mit S. degrangei, S. varie-gaturn, S. rheophilurn, O. ornater, O. spinosa, E. carthusiense, E. cryophilurn und S.reptans.
Th 5 bis Th 9: S. reptans, vergesellschaftet mit O. ornata, sporadisch mit W.equina sowie im Frühjahr mit Arten der beiden oberen Abschnitte.
Th 9 bis Th 10: S. reptans/ W. lineata, vergesellschaftet mit O. ornata, B. erythro-cephala, W. equina sowie im Frühjahr mit Arten der oberen Abschnitte.
Im obersten Abschnitt dominieren die montanen Arten, der zweite erhält durchAnteile campestrischer Arten submontanen Charakter. Auf den campestrischen drit-ten folgt ein potamo-campestrischer letzter Abschnitt. Es zei gt sich also eine guteÜbereinstimmun g mit den übri gen Proben der Langzeit- und der Querschnittsunter-suchung, welche der Assoziationsanalyse zugrunde lie gen. Eine permanente Zonie-rung lässt sich aber für die Thur nicht aufstellen, wie sie für die der Thur ähnlichenFulda in Deutschland anhand der Plecoptera (ILLIEs, 1953; P. ZWICK, 1969) und derSimuliiden (H. ZWICK, 1974) vorgeschlagen worden ist. Abgesehen von Abweichungenin den Faunenlisten fand ich in den unteren Abschnitten der Thur sowohl 1973, 1974wie auch 1975 Larven und Puppen der häufigeren montanen Arten P. rrJipes, S. varie-gatum, S. rheophilurn und der beiden Cleitosimulium-Arten, deren temporäres Vor-kommen im Frühjahr ich auf den fehlenden Temperaturgradienten sowie auf dieinfolge des stärkeren Gefälles höhere Drift (MOERGELI, 1974) zurückführe. Die Thurbildet im Gegensatz zur Fulda kein Krenal aus; die Rheokrenen auf dem Thurbodenwaren frei von Simuliiden und flossen so reichlich, dass schon nach etwa 50 m dieP. rufrpes-Populationen gefunden wurden. Die kleinen Rinnsale ihrerseits führen in-fol ge der starken Verkarstung des Gebietes nur temporär Wasser, während die Wild-hauser Thur, mit saisonbedingten Abwasserspitzen belastet, weitgehend simuliiden-frei ist. Somit wäre der ganze Thurlauf dem Rhithral zuzurechnen, das etwa bei Th 5in ein Epi- und ein Hyporhithral unterteilt werden könnte. Ab Th 9 treten in Gestaltvon W. lineata, B. erytlrrocephala und einem nach dem Zürcher Fischerei- nnd Pacht-verzeichnis gemischten Fischbestand potamale Elemente hinzu. Bei der Töss, ober-halb von Winterthur ein Forellen gewässer mit O. ornata und sporadischen P. hirtipes,E. vermin" und O. spinosa, markieren S. reptans, equina und gelegentliches Vor-kommen von W. lineata eine entsprechende fischereibiologische Grenze (vgl. auchRAI, 1970).
Um festzustellen, ob sich die Simuliidenarten anhand der Umweltfaktoren ähnlichgruppieren lassen wie anhand ihres Vorkommens, unterzog ich sie erneut einer Cluster-Analyse. Ich bezog alle Arten in die Untersuchung ein, für die ich über mindestenszehn Messwerte pro Umweltfaktor verfüge. Diesem Kriterium genügen die in Ta-belle 4 aufgeführten Arten, ausser O. spinosa. Als « Merkmale» dienten der Minimal-wert, der Maximalwert sowie der (verteilun gsunabhängige) Median jedes gemessenenMilieufaktors, nachdem die Daten der Lan gzeit- und der Querschnittsuntersuchungzusammen gelegt worden waren. Die Daten über Ma gnesium, Kalium, Natrium sowie
102 Vierteljahrsschrift der Naturforschenden Gesellschaft in Zürich 1978
den chemischen Sauerstoffverbrauch fielen we g. einerseits da fehlende Werte denRechenprozess beeinträchti gt hätten, anderseits da eine genügend en ge Korrelationzu anderen untersuchten Faktoren besteht. Somit standen 84 Masse zur Verfügung.Die ökolo gische Distanz d;,; wurde nach der bereits erläuterten Formel berechnet.Für die Clusterbildung wählte ich das UPGMA-Verfahren (Unweighted Pair GroupArithmetic Average Clustering). nachdem das Verfahren mit gewichtetem Mittelwert(WPGMA) nur unbedeutende Abweichungen ergeben hatte. Nähere Angaben, ins-besondere zur Erstellung und Interpretation der Phäno gramme, geben SOKAL undSNEATH (1963) und SNEATH und SOKAL (1973).
Das Dendrogramm für die 16 Arten (Abb. 4) gliedert sich in drei Gruppen, welchedie in der Assoziationsanalyse vorgeschlagene Gliederung teilweise mit gleicher Arten-zusammensetzung wiedererkennen lassen. Am en gsten schliessen sich die montanenArten zusammen, innerhalb derer sich die beiden alpinen Arten P. rufipes und E. car-thusiense, die sich bereits als si gnifikant assoziiert erwiesen haben (Abb. 3), von denanderen vier Arten ab grenzen lassen, die auch ins Alpenvorland hinab vorstossen.Die mittlere Gruppe umfasst die silvatischen und die sublacustrischen Arten. Die engeAssoziation der Berg- und Waldbachbewohner (Abb. 3) findet also bei Einbezug dergleich gewichteten Umweltfaktoren keine Entsprechung, was einerseits auf der stark
s.
U. U.0
Abb. 4. Phänogramm der 16 häufigsten Simuliidenarten der Schweiz, als Ergebnis der Cluster-Analysemit allen in der Lan gzeit- und in der Querschnittsuntersuchung erhobenen Daten.
Jahrgang 123 R. GLATTHAAR. Verbreitun g und Ökologie der Kriebelmücken 103
verschiedenen Höhenlage, Fliessgeschwindi gkeit, Turbulenz, Bachbettdimension,
Substrat sowie auf Unterschieden in eini gen chemischen Faktoren beruhen kann,
anderseits darauf, dass im Fall von E. cryophilum mehrere montane Fundstellen ein-
bezo gen worden sind. Die genannten Faktoren gleichen sich starker zwischen der
silvatischen und der sublacustrischen Gruppe als zwischen der montanen und der
silvatischen. Die Seeausflussbewohner spalten sich ihrerseits relativ früh auf, wohl
weil die Seeausflüsse besonders in physikalischer Hinsicht hetero gen sind. Die dritte
Gruppe des Dendrogramms zei gt den gewichti gsten Unterschied zur Assoziations-
analyse. Sie vereini gt die campestrische mit der potamalen Lebens gemeinschaft, wo-
bei 0. ornata und B. erythrocephala den beiden 117/hehnia-Arten und S. reptans ge gen-
überstehen, vermutlich weil beide auch langsamere, wärmere und chemisch stärkerbelastete Gewässer besiedeln.
Um Anhaltspunkte zu gewinnen, wie weit die physikalischen Umweltbedingungen
einerseits, die chemischen anderseits von Bedeutung sind, führte ich die Analyse je
einmal nur mit den physikalischen und nur mit den chemischen Daten durch. In bei-
den Dendrogrammen bleibt die Gruppe der sechs Bergbachbewohner erhalten, bei
den physikalischen Faktoren klarer als bei den chemischen. Auch die Untergruppe
P. rufipes ± E. carthusiense bleibt bestehen. Die mittlere Gruppe mit den silvatischen
und sublacustrischen Arten unterlie gt den stärksten Veränderun gen. Zwar bleiben die
beiden Waldbachbewohner E. vernum und E. COSICHUM durchwe gs beieinander, dies-
mal deutlicher bei den chemischen Faktoren. Die vier sublacustrischen Arten dagegen
schliessen sich zwar bei den physikalischen Faktoren locker der silvatischen Gruppe
an; bei den chemischen aber fällt E. angustipes heraus, vermutlich weil diese Art auch
in Gewässern mit höherer chemischer Belastun g vorkommt. Am schwierigsten zu
charakterisieren ist S. argyreatum, das sich wegen seiner ei genarti gen Spezialisierung
nur andeutun gsweise an die beiden anderen Seeausflussbewohner anschliesst. Die
dritte Gruppe mit den campestrischen und potamalen Arten bleibt, abgesehen von
leicht variierenden Verzwei gungssequenzen und -niveaus, durch alle vier Analysen
hindurch beisammen, am deutlichsten bei den chemischen Faktoren. Ledi glich bei
den physikalischen setzt sich B. erythrocephala etwas ab, was auf ihr Vorkommen in
extrem lan gsamen (bis wenige Zentimeter pro Sekunde), turbulenzarmen, tiefen und
im Sommer warm werdenden Gewässern zurückzuführen sein mag.
Die Analysen zei gen je nach dem zugrunde liegenden Material teilweise beträcht-
liche Abweichungen. Ich nehme an, dass das Dendrogramm nach den Umweltfak-
toren den wahren Verhältnissen am nächsten kommt, da eindeuti ge, spezifische An-
sprüche an einzelne Milieufaktoren auch bei nur zehn Messwerten im einschlägigen
Habitat zum Ausdrnck kommen. Anderseits fallen bei multifaktoriellen Analysen auf
einem oder wenigen Milieu-Elementen beruhende Biotopspezifitäten wie das reich-
liche Vorkommen von B. erythrocephala in langsamen Seeausflüssen zuweni g ins Ge-
wicht. Somit stellt sich bei weiter gehender Untersuchung die Fra ge nach der Gewich-
tung der einzelnen Komponenten, zumal da die durch geführten Analysen andeuten,
dass unter Berücksichtigung phänologisch bedingter Populationsdichte-Schwankun-
gen die physikalischen Faktoren einen relativ grossen Einfluss ausüben. Ferner ist es
wahrscheinlich, dass das Weibchen bei der Wahl des Eiablageplatzes auch durch
andere, hier nicht erfasste exogene und endo gene Faktoren beeinflusst wird.
104 Vierteljahrsschrift der Naturforschenden Gesellschaft in Zürich 1978
6. Autökologie der präimaginalen Kriebelmücken
Mit Hilfe der schrittweisen multiplen Regression ging ich die Frage an, in welchemMasse einzelne Umweltfaktoren die Verteilung und Abundanz der Simuliidenartenbeeinflussen. Sie gibt eine Reihenfolge der Faktoren nach Massgabe ihres Beitrages,den sie an den multiplen Regressionskoeffizienten R 2 leisten. Für die rechnerischeAuswertung benützte ich das Bibliotheksprogramm BMD 02R am Rechenzentrumder Universität Zürich. Für einige Proben führte das additiv-lineare Modell der Re-gression für die Populationsdichte, y, zu negativen Werten. Um diesen Fehler zukorrigieren, wurden diese Proben nebst eini gen «Ausreissern» eliminiert. Da auchProben mit kleinen positiven y-Werten vor allem aus phänologischen Gründen einensystematischen Fehler bewirken können, wiederholte ich die Analyse nur mit jenenProben, deren geschätzter y-Wert grösser ist als die Standardabweichung des Fehlers(STAHEL, mündl. Mitt.).
Mit Hilfe des Programmes BMD 05D liess ich für die häufigeren Arten die Streu-diagramme ausdrucken, in denen die Populationsdichte je Art als abhängige Variable(y, Ordinate) gegen die Messwerte jedes Umweltfaktors (x, Abszisse) aufgetragen ist.Faktoren, deren Werte über mehr als zwei Grössenordnungen in stark asymmetri-scher Verteilung variierten, wurden. wie in der Re gressionsrechnung, logarithmischtransformiert.
Obwohl die Arten nicht einheitlich reagieren, lassen die Streudia gramme funktio-
Legende zu Tabelle 6
Rangfolge der Umweltfaktoren nach Massgabe ihres Beitrages zuI «erklärten» Varianz (8 R 2 100)nach dem i-ten Rechenschritt der multiplen Regression. Faktoren, deren Beitrag <5% ist, sind ein-geklammert. Die Herkunft der Daten ist gekennzeichnet mit L, Lan gzeituntersuchung im KantonZürich, Q, Querschnittsuntersuchung der Schweizer Gewässer, I, Berechnung mit allen Proben, II,Nachrechnung nur mit den Proben, deren berechnetes y > s. Die zweitletzte Kolonne gibt denBeitrag des I. Faktors an, die letzte den Anteil der durch alle i über dem Einschlussniveau (F � 3)
liegenden Faktoren «erklärten» Varianz an. Die Nummern bezeichnen die Umweltfaktoren:
1 Höhe über Meer 17 Gesamthärte2 Wassertemperatur 18 Karbonathärte3 Flussbreite 19 Phosphatgehalt4 Flusstiefe 20 Nitritgehalt5 Fliessgeschwindigkeit 21 Nitratgehalt6 Abflussmenge 22 Ammoniumgehalt7 Turbulenz 23 Chloridgehalt8 Uferbeschattung 24 Gehalt an organischem9 Makrophyten im Wasser Kohlenstoff
S. variegatum Q 8 14 4 I (28) (7) (3) - - 7 7,8 29,2
S. rlreo ^kihuu iQ IQ II
1510
105
2929
27(7)
(5)(27)
--
-- -
--
--
55
8,99,6
30,831,4
S. argyreatuur L I 10 21 (19) (8) (3) (23) (II) (5) (9) 14 25,0 59,5
S. reptans (I
L 1L II
34
43
(13)9
(15)(13)
(9)(15)
(10)(24)
(I)(10)
(16)(6)
-(I)
-(23)
810
22,438,0
44,5 80,6
S. argerrtevstriahrni ' 1Q IQ II
730
(31)21
(19)14
(21)II
(I)(5)
(6)(23)
--
--
--
--
66
8,47,2
21,833,3
5(15
0:Pr00(Is.
aCD
CCDD'
0CD
CD
övI
106 Vierteljahrsschrift der Naturforschenden Gesellschaft in Zürich 1978
nelle Abhän gi gkeiten erkennen. Während die Messwerte y = 0 in fast allen Diagram-men über den gesamten Streubereich hinweg vorkommen, zei gen die Werte y>0 ver-schiedene Verteilungen. Eine breit streuende symmetrische Verteilun g bedeutet, inner-halb des Messbereiches, eine nur gerin ge Abhän gi gkeit. Solche Diagramme liefern inerster Linie das pH sowie andere nichttoxische hydrochemische Komponenten. Eineeng streuende symmetrische Verteilun g bedeutet modale Anhän gi gkeit; sie kommtvor beim Gehalt und Sätti gun gs grad des Sauerstoffs. Bei den übri gen Diagrammensind asymmetrische Verteilun gen die Re gel; sie deuten auf eine mehr oder wenigerstarke positive oder negative funktionelle Abhän gigkeit hin.
Tabelle 6 gibt die Ran gordnun g der Umweltfaktoren nach Mass gabe ihres Bei-trages, den sie an den Prozentsatz der «erklärten Varianz» leisten, der etwa zwischen20 und 80% liegt. Ein Vergleich mit den Streudia grammen zei gt, dass Faktoren mitstark asymmetrischer Verteilung die vorderen Rän ge bele gen. Ein Vorzeichentest mitden mittleren Rangzahlen für die ersten drei Rän ge je der physikalischen Faktoren(Nummern 1 bis 10) und der chemischen Faktoren zeigt mit einer Irrtumswahrschein-lichkeit von 5%, dass die physikalischen Komponenten mehr Gewicht haben in derRe gressions gleichun g als die chemischen, was auf einen stärkeren Einfluss auf Ver-teilung und Abundanz der präimaginalen Stadien bei den untersuchten Simuliiden-arten schliessen lässt.
Arten, bei denen der erstrangige Umweltfaktor einen relativ kleinen Teil derVarianz erklärt – eine willkürliche Grenze kann bei etwa 15% gezo gen werden – undbei denen mehrere Faktoren je einen Anteil >_ 5% erklären, können als euryök gelten.Dies ist der Fall bei O. ornata, S. variegation, S. rheophilurn und S. argenteostriaturn.Der relativ hohe Anteil des 1. Faktors in der Querschnittsanalyse dürfte auf den Ein-fluss der montanen Proben zurückzuführen sein, die in der Langzeituntersuchungfehlen. Deshalb scheinen aufgrund des «Datenmilieus» die genannten montanenArten eher als zu euryök, S. reptans dagegen eher als zu stenök bewertet. Ausserdemerreichen manche Faktoren mit stark linksschiefer Verteilung – die auch subjektivteilweise als wichtig eingeschätzt werden – das (willkürlich festgelegte) Einschluss-niveau (F > 3) nicht. Ebenso führte die Regression ohne die disqualifizierten Probennicht in allen Fällen zu subjektiv überzeugenden Resultaten.
Am stärksten überbewertet erscheint der Faktor Höhe bei S. argyreatum mit 25%Anteil (Tabelle 6), da die Art in ähnlicher Populationsdichte auch im Hahnensee-auslauf (804b) vorkommt, der mit 2150 m zu den höchstgelegenen Simuliiden-Fund-orten zählt.
Anhand der Streudiagramme und der besprochenen Analysen sei im fol genden dermögliche Einfluss einzelner Faktoren auf die Verteilung und Abundanz der Simu-lüden erörtert.
Die Höhe über Meer kann für das untersuchte Gebiet als wichti gster klima-bestimmender Faktor gelten. Sie beein flusst indirekt (über die Durchschnittstempe-ratur, Niederschla gsmen ge und die Dauer der Ve getationsperiode) Verteilung undAbundanz der Kriebelmücken. Sie ist mit verschiedenen Massen, die die Grösse unddie hydrochemische Belastun g der Gewässer anzei gen, ne gativ korreliert. Auf derAlpensüdseite wirken das grosse Gefälle und die kurzen Abflusswe ge den Folgengeringer Höhenla ge entgegen. So werden im Unterlauf des Ticino auch im Hochsom-
Jahrgang 123 R. GLATTHAAR. Verbreitung und Ökologie der Kriebelmücken 107
mer kaum je mehr als 15 bis 17° C gemessen. Wohl deshalb leben dort keine Flach-land-Simuliiden, dage gen werden oft Vertreter montaner Arten herabgeschwemmt.
Am stärksten sind, wie auch die multiple Regression (Tabelle 6) ergibt, P. rufipesund E. carthusiense an die Montanstufe gebunden. Nur im Flachland bis etwa 650 m
über Meer sind bis jetzt E. costatum, E. latigonium, die beiden Wilhelmia-Arten sowie
B. erythrocephala gefunden worden. O. ornata, E. vernum, E. angustipes, S. reptansund S. argyreatum entwickeln ihre Hauptpopulationen im Mittelland, reichen abermit einzelnen Ablegern in die alpine Region hinauf. So sind alle fünf Arten in densublacustrischen Biotopen des Oberengadins vertreten, denen die sich im Sommerrasch aufwärmenden Kleinseen zusammen mit dem geringen Gefälle ähnliche Be-dingungen verleihen, wie sie im Flachland herrschen. E. cryophilum und O. spinosascheinen in allen Höhen des Untersuchun gs gebietes (200 bis über 1500 m) heimischzu sein, wenn die Wassertemperatur nicht über etwa 17° C steigt, was im Mittellandnur in Waldbächen der Fall ist, in denen ich die beiden Arten gele gentlich gefun-
den habe.Weil die Wassertemperatur in allen untersuchten Gewässern im Winter knapp
über dem Gefrierpunkt liegt, dürfte der unterschiedlichen Sommertemperatur eineverteilungsbestimmende Rolle zufallen, und zwar um so eher, je länger sie wirkt.Erwartungsgemäss zeigen sich die Ber g- und. Waldbachbewohner kaltstenotherm; dieTemperatur ihres Milieus übersteigt selten 15° C. Die Flachlandformen ertragen etwashöhere Maxima, die im allgemeinen zwischen 18 und 23° C liegen. Auf 25° C undlokal sogar noch mehr können sich die meist an gut besonnten Orten lie genden Brut-
plätze von B. erythrocephala erwärmen (RUEHM, 1970). In stehendem Wasser undwährend des Transportes starben indessen die Larven bei Temperaturen über 25° Cbald ab, während die Puppen etwas resistenter waren.
Die die Flussgrösse umschreibenden Masse Breite, Tiefe und Abflussmengesind miteinander eng korreliert. Breite und Wassermen ge wirken wohl nur indirektauf die Kriebelmücken, nicht als einzeln fassbare kausale Faktoren. Mit zunehmenderBreite und Tiefe werden auch der Wasserkörper und das Lichtan gebot grösser, derAnteil der Uferpflanzen am Bewuchs jedoch kleiner, was vermutlich jene Arten be-günsti gt, die die Eier aus dem Flug ins Wasser werfen, zum Beispiel W. lineata, beider denn auch die Wassermenge als erster Faktor in die Regressionsgleichung tritt(Tabelle 6). Die Eusimulium-Arten und O. ornata zeigen in bezu g auf die Flussbreite
linksgehäufte, S. reptans dagegen rechtsgehäufte Streudiagramme.Eine Minimaltiefe scheint bei allen Simuliidenarten nur durch die Notwendigkeit
gegeben zu sein, dass der KöIper benetzt und die Prämandibularfächer mit demNahrungspartikel enthaltenden Wasserstrom versorgt werden müssen. Die grösstenTiefen wurden in potamalen Habitaten gemessen, mittlere in den campestrischen und
grösseren Bergbächen, während die Seeausflüsse 5 cm bis mehrere Meter tief seinkönnen. Waldbäche und überlaufene Dammkronen sind die im Untersuchungsgebietgefundenen Habitate mit Tiefen von weni gen Zentimetern, dafür relativ hohen Fliess-geschwindi gkeiten, welche im Verein mit dem Schatten beziehungsweise mit demgleichmässi g nachliefernden Reservoir des stehenden Gewässers die Simuliiden vorÜberwärmun g schützen. Anderseits konnte ich von der Elliker Fähre (Rh) aus mitdem Steuerhaken noch aus etwa drei Meter Tiefe Büschel von Ranunculus fluitans
108 Vierteljahrsschrlft der Naturforschenden Gesellschaft in Zürich 1978
und Potamogeton pectinatus erreichen, die mit Puppen von W. lineata und B. erythro-cephala besetzt waren. Eher stenotop sind die Prosimulium- und die Eusimulium-Arten.
Bei der Abflussmen ge sind wohl nicht so sehr die absoluten Werte massgebend,als vielmehr die Unterschiede in der Wasserführung zwischen Hoch- und Niedri g
-wasser. Diese betragen bei Seeausläufen nicht mehr als das Fünf- bis Zehnfache,während sie etwa in der Thur mehr als das Dreissi gfache erreichen können (MOER-GELI, 1974). Dieser Unterschied dürfte mit verantwortlich sein für das gehäufte Auf-treten von B. erythrocephala in Seeausflüssen vom Typ der Glatt bei Schwerzenbach(G 1) und ihr sporadisches Vorkommen in der fast so warm werdenden Thur (Th 9bis Th 10), wo sie sich auf relativ turbulenzarme Stellen beschränkt.
Turbulenz und Fliess g eschwindigkeit bilden in beiden Untersuchungenein eng korreliertes Faktorenpaar. Erwartungsgemäss ist die Turbulenz in den Ber g
-bächen am höchsten. Immerhin können grössere Flachlandflüsse wie die unteren Ab-schnitte der Töss, der Thur oder der Glatt während einer Hochwasserspitze vergleich-bare Werte erreichen. Waldbäche und noch mehr die Seeausläufe, zu denen auch derHochrhein zu rechnen ist (DEMMERLE, 1966), unterliegen diesen Veränderungen inabgeschwächtem Mass und mit grösserer zeitlicher Verzö gerung. An Ausläufen, woS. argyreatum zur Massenentwicklung kommt, herrscht entweder eine rasche, relativwenig turbulente, oder aber eine relativ langsame, stark turbulente Strömung vor. ImGebirge sind die Ansprüche dieser Art weniger spezifisch.
Obwohl eine minimale Fliess geschwindigkeit für alle Arten lebensnotwendig ist,steht sie in der multiplen Regression nirgends an erster Stelle. Zwar lassen sich Krie-belmtickenlarven in einer Petrischale mit etwa 4 mm Wasser bei 4° C gut zwei Wochenlan g am Leben erhalten, nicht aber zur Verpuppung brin gen. Oli gorheobiont sind dieSeeausflussbewohner B. erythrocephala und E. angustipes, an deren Brutplätzen dieFliess geschwindigkeit 10 bis 40 cm/sec beträgt. Als langsamstes kriebelmückenhaltigesGewässer erwies sich der Hasenseeauslauf mit 0 bis 10 cm/sec, wo Larven und Puppenvon B. erythrocephala noch an Stellen mit 1 cm/sec zu finden waren. Das Optimumaller übri gen Arten liegt bei 0,5 m/sec oder darüber, am höchsten bei P. rufipes, E. car-thusiense, S. rheophilurrr und S. argenteostriatum.
Die Uferbeschattun g kommt wohl bei E. costatum als direkt die Biotopwahldes Weibchens beeinflussender Faktor in Fra ge, bei den übri gen Spezies eher alsindirekter (Schutz vor Überwärmung). Wo ein Waldbach in offenes Gelände hinaus-tritt, löst innerhalb weni ger Dutzend Meter O. ornata die silvatische Simuliidenfaunaab; für E. costatum erwies sich die Grenze in allen Fällen als scharf, während E. ver-num auch in gut belichteten, oligosaproben Gewässern in grösserer Zahl gefundenwurde, welche aus klimatischen (Auslauf des Lej Nair, 804a) oder hydrologischenGründen (Grundwasserkanal, GK) nicht wärmer als 17 bis 20° C werden. Die Artwird auch gelegentlich verdriftet, wie Einzelfunde im Schneitbach (Sn) und 1976 inder Reppisch zeigen. Die Brutplätze der übrigen Arten befinden sich in offenem bishalbschattigem Gelände, zumal da die Ima gines ta gaktiv und die Larven positivphototropistisch sind (GRENZER, 1949, und eigene Beobachtungen). In Wiesenbach-quellen leben neben O. ornata und E. vernurrr Vertreter der Gruppe E. aureum undE. angustitarse. Bemerkenswert ist die Massenentfaltung von S. argyreatum-P opu-
Jahrgang 123 R. GLATTHAAR. Verbreitung und Ökolo gie der Kriebelmücken 109
lationen in dunklen Abflussschächten (ZWICK und RUEHM, 1972), die ich selbst imAusflussschacht des Lucelle-Teiches (127) beobachtete.
Als Substrat, auf dem sich die halbsessilen Präima ginalstadien festheften, kom-men verschiedene Materialien in Betracht, sofern ihre Oberfläche dem Abdomenendeder Larve oder der Sohle der Puppe ein Anheften ermöglicht und sofern es genügendverankert ist, um der Strömun g standzuhalten. Das feinste Substrat, auf dem ichSimuliiden gefunden habe, sind Cladophora-Büschel, in denen ich Junglarven vonO. ornata fand, nicht aber Altlarven und Puppen. Mit Sphaerotihrs, schleimi gen Algen(insbesondere Diatomeen) oder Schlick überzogene Steine erwiesen sich stets als krie-belmückenfrei. Stehen sowohl Makrophyten wie auch Steine als geeignetes Substratzur Verfü gung, sind die Pflanzenblätter in der Re gel dichter besiedelt als die Steine.Nur S. argenteostriatum und S. degrangei sassen häufiger an Steinen und Ästen alsauf dünnen Zwei gen und Blättern, die von den anderen montanen Arten gelegentlichdicht besiedelt sind. Im Unterlauf der Thur (Th 8 und 10) entwickeln S. reptans undW. lineata ihre Frühjahrspopulationen auf den vom Ufer ins Wasser ragenden Salix-
Zweigen und -Blättern. Erst die Sommer- und Herbstgeneration besiedelte auch dieGerölle des Flussbettes, auf denen die Präima gines im Sommer 1973 dichte Überzügebildeten. Als oft dichtestbesetzte Substrate erwiesen sich in den untersuchten Habi-taten Folien und andere Abfälle aus Plastik; der Dichteunterschied zu anderen Sub-straten der Umgebung betrug oft mehr als eine Zehnerpotenz.
Lediglich O. ornata, B. erythrocephala, E. latigonium und die E. aureurn-Gruppedürften für eine erfolgreiche Fortpflanzung auf benetzte, an der Wasseroberflächeschwimmende Pflanzenteile angewiesen sein. Diese Arten entwickeln nur in den mitMakrophyten bewachsenen Bächen grössere Populationen, wie auch die Regressions-rechnun g andeutet (Tabelle 6). Bei der Tösse gg (Tö 3) fand ich Laichschnüre vonS. reptans, die locker an Salix-Zwei gen klebten. Die beiden 1J'ilhelnria-Arten werfendie Eier aus dem flussaufwärts gerichteten Flu g ins Wasser. Nur gelegentlich findetman die unregelmässi gen Muster ihrer Gele ge an benetzten Pflanzen (RtiEHM, 1971,und ei gene Beobachtungen). Im Potamal der Glatt und des Hochrheins nehmen diereichlich wuchernden Ranunculus fluitans-, Potamogeton pectinatus- und Myriophyllu n-Arten eine dichte Simuliidenpopulation auf und ermöglichen trotz fehlenden oderstark veral gten und verschlickten Steinen eine Massenentwicklung.
In der Kempt (Ke) beobachtete ich, dass die Cladophora-Massenentwicklung desSommers 1974 mit einer gegenüber dem Vorjahr auf 10 bis 20% reduzierten Simu-liidenfauna einher ging. Diey Al ge deckte über 90% der Wasserfläche, bremste denFluss stellenweise und deckte auch als Eiabla geplatz geei gnetes Substrat zu. Diesbetrachte ich als direkte Ursache des Populationsdichte-Rückganges von O. ornata,da sich die Gewässerqualität in beiden Jahren entsprochen hat.
Von den silvatischen Arten ist anzunehmen, dass sie ihre Eier an verschiedenartigesSubstrat heften, zum Beispiel auch unmittelbar an den Austritt von Rheokrenen, woich ihre Larven schon gefunden habe.
Die Sauerstoffs ättigun g lieat in den weitaus meisten Simuliidenhabitaten nahebei 100%, meist etwas darüber. Bisher fand ich nur B. erythrocephala, W. equina undO. ornata in sauerstoffarmem und zugleich stark verschmutztem Wasser, beispiels-weise unterhalb von Abwasser-Einläufen. Besonders eurytop verhält sich B. erythro-
110 Vierteljahrsschrift der Naturforschenden Gesellschaft in Zürich 1978
cephala, die einerseits im stark abwasserbelasteten Furtbach (416) bei nur 35%, ander-
seits in der frühsommerlichen Glatt (G 1) bei etwa 320% Sauerstoffsätti gung gefundenworden ist. Diese Übersätti gung, eine Folge der Al genblüte im stark eutrophiertenGreifensee, war bei G 2 auf 160% abgebaut. Bei den von S. argyreatum besiedeltenAusflüssen la gen die Extremwerte näher beisammen, ebenso an den Fundorten dersublacustrischen Eusimulium-Arten. welche oft erst eine gewisse Strecke unterhalb desAuslaufes vorkommen. Dass die extremen Sauerstoffverhältnisse das Fehlen vonS. argyreatum am Re gulierwehr des Greifenseeausflusses mitbe gründen, ist nicht aus-zuschliessen.
Als Masse für die or g anische Belastung der Gewässer standen der biochemi-
sche Sauerstoffverbrauch nach 5 Tagen, der chemische Sauerstoffverbrauch sowie derGehalt an or ganischem Kohlenstoff zur Verfü g ung, die untereinander in gewissemMasse korreliert sind. Die gemessenen Werte lie gen in den Ber g- und Waldbächenam tiefsten. Eine relativ gerin ge Streuung wurde auch in den sublacustrischen Habi-
taten nachgewiesen, während die Spitzenwerte von den stark belasteten Stellen des
Furtbaches (416), des Altbaches (Al). der Glatt (G 2ff.) und der Töss (Tö 2 und 3)sta mmen. Auch das durch Hochwasser in Bewe g un g gebrachte Sediment liefertegelegentlich hohe Kohlenstoffwerte.
Das pH zeigt bei den Streudiagrammen am häufigsten eine breite, symmetrischeStreuun g und tritt dementsprechend selten in die Regressions gleichun g ein. Somitlässt sich aufgrund der vorliegenden Messwerte bei keinem Habitat eine Abhän g i g
-keit der Simuliidenfauna von der Reaktion des Mediums erkennen. pH-Werte unter 7stammen vornehmlich aus Proben mit niedriger Härte und Leitfähigkeit und betreffen
vor allem die montanen und alpinen Arten. GRUNEWALD (1973) gelang indessen dieZucht von B. ervthrocephala auch bei pH-Werten unter 7. Die höchsten Werte (über
9) wurden im Frühsommer infolge der C09-Verarmnng durch die Assimilationstäti g-keit der Algen erreicht.
Die L e i t fä h i g k e i t des Wassers entsteht durch dessen Gehalt an gelösten Ionen,von denen Calcium, Magnesium. Karbonate und Sulfate den Hauptanteil stellen.
Diese Proportionalität schlägt sich auch in engen Korrelationen nieder, die eine sum-
marische Besprechun g erlauben. Dieser Faktorenkomplex, der bei den Bergbächen
die grösste Streubreite zeigt – von 20 bis über 1500 Mikrosiemens –. liefert keine
Anhaltspunkte für eine Beeinflussung der Verteilung und Abundanz der Simuliiden.
Dagegen kann ein Einfluss von seiten der anionischen Belastungsanzei ger (Stickstoff-verbindun gen, Phosphat, Chlorid) bestehen; von Ammoniak und von Nitrit ist be-
kannt, dass sie auf Vertebraten. beispielsweise Elritzen, toxisch wirken können, Am-
moniak besonders bei hohen pH-Werten und hohen Temperaturen (HARNISCH, 1951;
MERKENS und DOWNING, 1957). Berg- und Waldbäche liefern die niedri gsten Titer,gefolgt von den Seeausflüssen, während die potamalen und die campestrischen Habi-
tate am stärksten belastet sind. Nur bei O. ornata treten in der Re gression Belastun gs-anzeiger in positive Korrelation zur Populationsdichte.
Der Fi1terriicl stand enthält unter anderem die den Kriebelmückenlarven direkt
zur Nahrung dienenden Stoffe. In den Ausflüssen eutropher Seen ist er im Sommerinfolge der Al genblüte grün, in den übrigen Tieflandbächen und -flüssen tritt er in
verschiedenen Brauntönen auf, während der Rückstand der Bergbäche grau erscheint.
R. GLATTHAAR. Verbreitun g und Ökolo g ie der Kriebelmücken 111Jahrgang 123
Nach unten scheinen die gemessenen Werte des Schwebestoffgehaltes nir gendwo be-grenzend zu wirken, lie gen doch auch die Mittelwerte fast durchwe gs unter 10 mg/Liter. Eine im Kanton Zürich bei Hochwasser genommene Probenserie zeigt Extrem-werte, die mehrere Gramm pro Liter erreichen. Die spärliche Benthosfauna in «Glet-schermilch» führenden Bergbächen gegenüber solchen mit klarem Wasser und sonstähnlichen Bedingun gen deutet darauf hin, dass der hohe Anteil an unverdaulichemSilt ein Hindernis für eine dichtere Besiedlun g darstellt.
In der Absicht, weitere Hinweise auf die Einwirkung von Umweltfaktoren auf dieSimuliidenfauna zu erhalten, unterzog ich auch die Probestellen anhand der gemes-senen Faktoren einer Cluster-Analyse, nach dem besprochenen Verfahren je einmalmit allen, nnr mit den physikalischen und nur mit den chemischen Faktoren. DasPhäno gramm der Zürcher Gewässer mit allen Umweltfaktoren (Abb. 5) zeigt, aufdem Niveau von 1,5 geschnitten, vier Gruppen und einen Einzelgänger. An diesenProbestellen kamen mit Ausnahme der Thur (Th 10) nur nichtmontane Simuliiden-arten der Assoziationen I1 bis V vor. Die Funde der silvatischen Arten sind im oberenTeil der zweiten Gruppe integriert, der die wenig belasteten, quell- und grundwasser-haltigen Feldbäche umfasst. Der andere, grössere Teil dieser Grnppe führt die stärkerbelasteten campestrischen Bäche. Ausserhalb derselbeH sind von O. ornata besiedeltdie Bäche der ersten Gruppe: der Altbach, der wohl infol ge seiner hohen chemischenBelastun g mit der Glatt (G 2 bis G 5) assoziiert ist, ferner der Schneitbach (3. Gruppe)sowie sporadisch der Rhein und die Thur (4. Gruppe). Gut miteinander verbundensind die beiden kleineren, turbulenten Teichausläufe (Lützelsee, Walcheweiher), wäh-
Abb. 5. Phänogramm der 25 Probestetlen der Lan gzeitbeobachtung. Abkürzun gen wie in Abb. 2.Cluster-Analyse, durchgeführt mit allen Umweltfaktoren.
112 Vierteljahrsschrift der Naturforschenden Gesellschaft in Zürich 1978
rend G 1 wegen verschiedener physikalischer wie auch chemischer BesonderheitenEinzelgän ger ist. Ähnlich wie die sublacustrischen werden auch die potamalen Habi-tate auseinandergerissen, wohl vor allem deshalb, weil der Rhein bedeutend grösserist als die Glatt, in zweiter Linie, weil er zivilisatorisch nicht so stark belastet ist wiediese. Bei der Töss nnd bei der Glatt schliesslich gibt die Analyse mit den physika-lischen Faktoren die biozönotische Gliederung wieder. Bei der Glatt ist diese in mehr-facher Hinsicht ungewöhnlich. Der Greifenseeausfluss (G 1) bildet bei nur sechs bissieben Meter Breite ein zunächst sublacustrisches Potamal aus, mit einem reichenBewuchs an Phragrnites, P/ralaris und anderen hydrophilen Gramineen. Etwa dreiKilometer weiter unten werden auch submerse Makrophyten häufiger: Myriophyl-lurn, Ranunculus, Potamogeton. Bei ständi g steigender chemischer Belastung bleibendie physikalischen Verhältnisse bis Hochfelden (etwa 3 km unterhalb Niederglatt,G 4) ähnlich, bis zum Stau des alten Elektrizitätswerkes Glattfelden. Der untersteAbschnitt, G 5, nimmt da gegen rein rhithralen Charakter an. Mit dem Gefälle, dasvon 2,5 auf 7 Promille steigt, nimmt auch die Fliessgeschwindi gkeit und Turbulenzzu und die Tiefe, die Temperatur und der Makrophytenbewuchs ab, während sich derChemismus trotz etwas Grundwasserzufluss (WASER et al., 1934) gegenüber G 4 nurgeringfügig ändert.
Während im sublacustrischen Habitat bei Schwerzenbach (G 1) über die ganzeBeobachtun gsdauer sowie in späteren Proben nur B. er.vtlurocephala nachzuweisenwar, setzte oberhalb der Aubrugg (G 2) eine stärkere W. lineata-Population ein, be-gleitet von etwas W. equina (Tabelle 7). Bei G 3 und G 4 führte die Glatt, wohl wegender lokal etwas erhöhten Turbulenz und Geschwindi gkeit an Prallhän gen, auch O.oruata, die ihren Beitra g an die erhöhte Diversität dieser Probestellen leistete. BeiGlattfelden (G 5) hingegen dominierte O. oruata, gefolgt von Ir. eguina, währendB. er_vthroceplrala und W. lineata offenbar hier nicht zur Fortpflanzung kamen. Dievereinzelt gefundenen Individuen dieser Arten dürften mit der Abdrift aus dem oben-lie genden Potamal zu erklären sein. Es zei gt sich somit bei der Glatt besonders deut-
Tabelle 7. Kriebelmückenfunde in verschiedenen Abschnitten der Glatt, Anzaht Larven und Puppen.Die Probestellen sind wie in Tabelle 1 bezeichnet, die Zeit ist in Monat und Jahr angegeben. H: Diver-
sitätsindex nach SHANN0N/WIENER
Cluster-Analysenach Arten mit allen
Umweltfaktoren
Cluster-Analysealler Probestellen mit allen
Umweltfaktoren
Multi-dimensionalScaling
Angaben von ZWICK(1974)
potamale Gruppe
canlpestrische Gruppe
montane Gruppesensu strict ()
submontane Gruppe
silvatische Gruppe
potanlo-cam pest rischeGruppe
montane Gruppesensu strict()
submontane Gruppe
silvatische «Gruppe»(nur I Probestelle)
grosse, wenig belasteteGewässer
kleinere, stark belasteteGewässer
O.-arnala-Biiche
O.-ornata-/S.-replans-13iiche und -Müsse
montane Gruppe
silvatische Gruppe
potamale Gruppe
campestrische Gruppe
montane Arten
rhithrale Arten
silvatische Arten
potamale Artenl
hyporhithrale Arten
sublacustrische Gruppe sublacustrisehe Gruppe sublacustrische Gruppe sublacustrische Arten
Speziesgruppennach der Assoziations-
Analyse
montane Gruppe
silvatische Gruppe
Tabelle B. Gegenüberstellung der Ergebnisse der Assoziationsanalyse und der ökologischen Analysen. Punktierte Abgrenzung bedeutet Wechset-beziehungen zwischen Gruppen, leeres Feld bedeutet andersartige Gruppierung der Arten oder Gewässer
Zusammensetzung in der Reihenfolge abnehmender Fliiuligkcit: IV. equina, IV. lineala, O. ornara, 13. eri'lhroeephala.
w
114 Vierteljahrsschrift der Naturforschenden Gesellschaft in Zürich 1978
lic6, dass die Simuliidenfauna weniger dem Chemismus als dem inversen pbysiba|isobon Gradienten fold. Die Fische verhalten sich ähnlich; die Glattfelder Reviereim alten Bachbett gelten als Furc||cogewüsscr, während das Zürcher Pachtverzeichnisfür {] 1 bb G 4 gemischten Fischbestand angibt. 'Äbn|icbc Vcrbü\toioso scheinen im8üttwücmeehucb(8ö) vorzulie gen, dem ich im Sommer 1973 uOc50Om6ne Probeentnahm.
Eine Cluster-Analyse, die ich mit den 85 faunistisch er giebi gsten Proben der Quer-schnittsuntersuchung durchführte, er gab ebenfalls eine grössere Anlehnun g der Ver-trüungdec8imulüdeourteuuodiopbysikaÜsc6enFaktorcoa\oaudiechomischeo.
Bei vergleichender Betrachtung der Gewässerdendrogramme zei gt sich, dass dieAufgliederung um so feiner ist,jc mehr einander ähnliche Proben zur Verfügungstehen. Deshalb unterteilt die Analyse der Zürcher Gewässer die campestrischen, dieAnalyse der Schweizer Gewässer die montanen Habitate. Stellt man alle vier bespro-cbrncn Cluster-Analysen nebeneinander der Assoziationsanalyse gegenüber, wie dieTabelle 8 zei gt, so findet diese aerudc durch jene Analysen ihre 0cstÜd guog, die fürdie betreffende Arten gruppe am kompetentesten sind. Die Lan gzeituntersuchung gibtin der Cluster-Analyse die campestrische und soweit möglich auch die potamale unddie silvatische Gruppe wieder, die Querschnittsanalyse die montane und die cam-pestrische. Das Phänogramm der Arten nach Umweltfaktoren gibt die Assoziationenamgctrcucstenvicdcr;cskuno|odiglicbdiupotamalcuunddicoumpegdscbcoArtroaufgrund der ein ge gebenen Daten nicht voneinander trennen. Schwächer ist die sub-lucustösc6eGruppcfüodier1.dabciibrf\usnub\uodGuv/icbcungdcrFuktoreouocbproblematischer sind und diese Habitate in bezu g au[ die erfassten Faktoren starkvariieren. Auch die autökologischen An gaben von Zw/cx (1074) zu den einzelnenArten passen mit Ausnahme der im Kanton Zürich als sublacustrisch charakterisier-
7. Prognostische Bedeutung der Analysen
NicbtuUcdcrbydrocbenuischuotenucb»rnGcvüsscrnurenzurUntcrsocbuogaznitvonSimu|iidcnbcsicdclt.Dmzuc,bzbreo.obdicaogcnoodteoMetbodcnvnoprogoo'stischem Wert sind, bezo g ich bei der Cluster-Analyse der Zürcher Gewässer zwei, beider Cluster-Analyse von 100 Proben aus der übrigen Schweiz 15 Fundstellen ohneKricbc|mücüzn ein.
Auch die multiple Regression erwies sich als prognostisch verwertbar. Für jedeProbe und jede untersuchte Spezies ver glich ich die berechnete Populationsdichte, y'mit der tatsächlich gefundenen sowie mit dem Betrag der Standardabweichung svooK,v/o6riidcoIodcxdes|ctztcnFuktoobczeicbuu.dernoücincozF'Wnrt 3,0in die Gleichun g einbezo gen worden ist. Lie gen die errechnete und die beobachtetePopulationsdichte 6ncr Probe Ober dicserGroozc ' sprechen wir von «re gulärem Vor-kommen», lie gen beide darunter, von "rrgu|ürem Fe@cn^ der betreffenden Simu'liidooart an der untersuchten Probcstelic Lie gt die gefundene Dichte Jarübcr, dieberechnete darunter, vermerken wir «paradoxes Yorbommcnx, im reziproken Fall«erwartetes Vorkommen» ~~xparudoxcs Fehlen».
Stellt man für die kriebelmückenfreien Probestellen die von der multiplen Regres-
Jahrgang 123
R. GLATTHAAR. Verbreitung und Ökolo g ie der Kriebelmücken 115
sion aus zu erwartenden Spezies den Arten gegenüber, die bei der Cluster-Analyse fürdie Schweizer Gewässer in der entsprechenden Cluster-Gruppe am häufigsten nach-gewiesen worden sind, so ergibt sich eine gute Übereinstimmung, wobei die Angabender multiplen Regression etwas differenzierter sind. Bei den simuliidenfreien Probe-stellen handelt es sich meist um grössere, wasserreiche Alpenflüsse, die im Sommeroft trübes Hochwasser führen, so dass die Proben unter erschwerten Bedingungengenommen werden mussten. Die Stellen liegen an der Rhone, der Lütschine, der Aareund im En gadin; in wechselnder Frequenz oder Hierarchie «sollten» die sechs mon-tanen Arten vorkommen; im Tessin, im Scairolo bei Fi gino und im Laveggio beiCapolago, werden O. ornata und S. variegation erwartet. Für die Brauchbarkeit der-arti ger Prognosen sprechen verschiedene Funde, die nach 1974 gemacht worden sind:
— Anfang September 1975 wurden im Flazbach bei Samedan die «erwarteten» ArtenP. rufipes und E. carthusiense gefunden.
— Im Sommer 1975 wurde im Doubs bei St-Ursanne (509) neben S. reptans und O. or-nata die «vorhergesa gte» W. lineata gefunden.
— Am 31. Mai 1976 fand ich im Zürcher Binnenkanal (ZB) E. vermin?, das in allenneun Proben der vorhergehenden Jahre «erwartet» worden war.
— Anfang Juli 1977 konnte ich im bisher simuliidenfreien Chriesbach (Ch) erstmalseinen reichlichen Besatz an Kriebelmücken nachweisen. Es handelt sich um diesowohl von der Cluster-Analyse wie auch von der Re gression her «erwartete» ArtO. ornata, deren Populationsdichte jener der in bezug auf die Umweltfaktoren ähn-lichsten Bäche (Kempt, Näfbach. Abb. 5) entspricht. Nachdem die Kolonisierung(oder Rekolonisierung?) des Chriesbaches durch O. ornata dem Abschluss deroberhalb der Probestelle betriebenen Bautätigkeit folgte, vermute ich im früheroftmaligen Auftreten und im gegenwärtigen Ausbleiben der Schübe erdiger Sus-pension die Ursache für das beobachtete Verhalten von O. ornata.
In der anderen simuliidenfreien Probestelle der Langzeituntersuchung, dem Wal-cheweiher-Auslauf, wird S. argyreaturn «erwartet». Ein Gelege der Art, das ich imAugust 1974 dort einsetzte, lieferte zwar Larven und schlüpfende Puppen, aber keineNachkommenschaft, wohl wegen zu geringer Wassermenge und zu starkem Algen-wachstum (Spirogyra).
Schliesslich ist bemerkenswert, dass die multiple Regression die Schwarze Lüt-schine unterhalb von Lütschenthal und die Rhone bei Chalais für keine der unter-suchten Arten als passenden Habitat bewertet. An diesen Probestellen wies auch dieBegleitfauna eine starke Verarmung auf; in diesen ständig von anorganischem Siltgetrübten Gewässern fanden wir ledi glich einzelne Chironomiden-, Baetiden- undBlepharoceridenlarven. Anderseits werden besonders in der Querschnittsuntersuchungdie vier montanen Simuliidenarten relativ oft als «paradox vorkommend» gemeldet,so in der Ri gi-Aa bei Arth (308) E. cryophilum und S. variegation, oder in der KleinenEmme bei Blatten LU (318) S. rheophilum und S. argenteostriaturn, wo nur O. ornataregulär vorkommt. Ein Blick auf die Stelle oberhalb der Fontannen-Mündun g (314),wo alle vier Arten regulär leben, zeigt, dass die Art des Einzugsgebietes (Flachland,Gebir ge oder See) und die Zusammensetzung seiner Fauna mass gebende Faktorendarstellen, die bei der Analyse eines kleinen Flussabschnittes zu kurz kommen.
116 Vierteljahrsschrift der Naturforschenden Gesellschaft in Zürich 1978
B. Diskussion
Während die Fauna der grösseren Fliessgewässer vor allem durch die Querschnitts-untersuchung als relativ gut erfasst gelten kann, bestehen bei den zahlreichen kleinenund kleinsten Gewässern, vor allem in der alpinen Quellregion, noch grosse Lücken,wie zum Beispiel der erst 1977 geführte Nachweis von E. crenobium zeigt. Somit dürftedie Nachweisdichte vor allem bei T. Irydroides, den Prosimulium- und Eusimulium-Arten bei weiterführender chorologischer Arbeit anstei gen. Auch für T. bezzii sind inder Westschweiz weitere Fundnachweise zu erwarten.
In allen Nachbarländern sind längere Artenlisten aufgestellt worden. GRENIER
(1953) führt flir Frankreich 30 Arten an (einschliesslich einige Unterarten, die heuteals Arten gelten), ZWICK (1974) für die Bundesrepublik Deutschland 31 Arten, ZWICK
(1976) für Österreich 30 und schliesslich RIVOSECCHI und CARDINAL' (1975) für Italiennicht weniger als 89 Spezies. Für die Schweiz dürften etwa dieselben rund 50 Artenzu erwarten sein, die KNOZ (1965) für die Tschechoslowakei an gibt. ZWICK (1976)schätzt für Österreich dieselbe Artenzahl.
Aus allen Nachbarländern gemeldet werden und deshalb auch in der Schweiz zuerwarten sind P. tomosvar vi, E. aw eum und Obuchovia auricoma; ob zwei Puppen-kokons mit hohem Kragen, die ich im September 1974 in der Rigi-Aa bei Arth (308)fand, zur letztgenannten Art gehören, ist nicht zu entscheiden (ZwtcK, in litt.). Weiterein Nachbarländern gemeldete Arten sind E. carpathicum, S. austeni (= venustum),S. verecundurn (= sublacustre), E. truncation (Zwick, in litt.), Schönbaueria pusilla,S. galeratum, S. morsitans nnd Vertreter der Gattun g Cnephia (GRENIER, 1953).
Da die Areale aller in der Schweiz nachgewiesenen Simuliidenarten sich zum grös-seren Teil auf das Ausland erstrecken, ist damit zu rechnen, dass in der vorliegendenUntersuchung nicht einmal alle Habitattypen einer Art erfasst worden sind. Als Bei-spiel dient eine Gegenüberstellun g meiner Daten für O. ornata mit jenen, die RuEH'vl
(in litt.) an der Jürse, einem Moor gewässer, erhoben hat, wo die Art vorkommt (Ta-belle 9). Dieser Habitat des eisenhaltigen und wohl deshalb auch sauern und karbonat-armen Moorbaches fehlt in den Schweizer Proben. Ebenso verfügt GRUNEWALD (1972,1973) über Brutnachweise von B. erythrocephala in weicherem Wasser, wogegen dieschweizerischen oft in härterem und stärker belastetem Milieu lie gen (Tabelle 10).
Der Diskussion über den Einfluss bestimmter Umweltfaktoren auf Verbreitungund Abundanz einzelner Simuliidenarten fehlt auch der Überblick über die zeitlicheDimension. Erste numerische Angaben über das Milieu der Präimaginalstadien stam-men von GRENIER (1949): Sauerstoffgehalt, pH, Calciumhärte; weitere von CHUTTER(1968) und MUIRHEAD-THOMSON (1970): Gesamthärte und freies Chlorgas im Zucht-wasser, sowie am ausführlichsten von CARLSSON (1962. 1968), welcher nicht nur hydro-chemische Faktoren analysierte, sondern auch mehrere physikalische Umweltfaktorenprotokollierte: Art des Substrates. Fliess geschwindigkeit, Höhe über Meer, Fluss-breite, ferner, allerdings mehr qualitativ und summarisch, Licht- und Nahrun gs-an gebot sowie die Belastung, «pollution». Erst mit GRUNEWALDS (1972, 1973) Unter-suchung an B. erythrocephala liegt ein genü gend grosses und vergleichbares Daten-material über 18 Umweltfaktoren vor, das weni gstens für diese eine Spezies Rück-schlüsse auf die Ökologie und Physiologie der Präimaginalstadien ermöglicht.
Jahrgang 123
R. GLATTHAAR. Verbreitung und Ökolo gie der Kriebelmücken 117
Tabelle 9. Umweltdaten für O. ornata aus der Jürse (RUEHM. 1974, in litt.) und nach eigenen Mes-sungen. Die Härten sind auf französische Härtegrade umgerechnet
1 Nach MOERGELI (1974) für die Thur und ihr Einzugsgebiet.
Tabelle 10. Milieufaktoren für B. eryrhrocepliala, die sowohl von GRUNEWALD (1972, 1973) wie auchin der Schweiz gemessen wurden. GRUNEWALDS Daten sind, soweit erforderlich, umgerechnet worden.
= arithmetischer Mittelwert, Min. = kleinster, Max. = g rösster Messwert, n = Anzahl Proben
1 Nur eigene Daten (n = 70). 2 Nur Querschnittsdaten (n = 12).
Obgleich in den Zürcher Gewässern schon seit einigen Jahrzehnten gründlicheUntersuchungen über hydrochemische Faktoren durch geführt werden (WASER et al.,1934, 1944; DEMMERLE, 1966; FLEISCH, 1970; MOERGELI, 1974; RAI, 1970; ZOBRISTet al., 1976; FERRET, 1977) und die daraus ersichtliche Eutrophierung hinlänglich
118 Vierteljahrsschrift der Naturforschenden Gesellschaft in Zürich 1978
bekannt ist, lässt sich infolge fehlender Faunenanalysen nichts aussagen über den Ein-fluss dieser Veränderungen auf die Kriebelmückenfauna.
Mit nur punktuellen Wasseranalysen werden kurzfristige periodische Schwankun-gen wie zirkadiane Abwasserschübe ebensowenig erfasst wie die relativ seltenen, aberfür die Fauna oft verderblichen Giftstoff-Unfälle. Die Verluste an Kriebelmückenwerden aber offensichtlich durch die Drift bald wieder kompensiert. GRUNEWALD
(1972, 1973) und RuEHM (1970a) messen der zirkadianen Temperaturschwankung, diegerade in den langsamen, oft auch untiefen B.-erythrocephala-Gewässern bis über5° C betra gen kann, eine gewisse Bedeutun g als Entwicklungsmotor bei. Aber auchin schneller fl iessenden Gewässern erreicht diese Amplitude noch beachtliche Werte,wie aus den Messungen von MOERGELI (unpubliziert) hervorgeht, der bei Altikon(Th 10) je einmal im November 1969 und im Juni 1970 über 24 Stunden hinweg allezwei Stunden eine Probe nahm. Besonders bemerkenswert sind die Temperaturampli-tuden (Juni: 19,0/23,4° C, November: 6,2/9,8° C), der Sauerstoffgehalt mit einem Mit-tagsmaximum von 15,5 mg pro Liter im November und 8,75 mg/Liter im Juni; dienächtlichen Minima lagen bei 7,7 beziehungsweise 5,45 mg/Liter. Im gleichen Sinnevariierte die Sauerstoffsättigung zwischen 61 und 138% im November und zwischen60,8 und 100% im Juni. Diese Schwankungen können auch natürlich bedingt sein.Eine schwache Periodik zeigten der Kaliumpermanganatverbrauch, der Ammonium-gehalt, der BSB5 und der Nitratgehalt, während die Härte und der Phosphatgehaltso gut wie konstant blieben.
Immerhin können die vorliegenden Daten und Auswertun gsmethoden doch als einbrauchbarer autökologischer Ansatz für die Untersuchung der Lebensbedingungenmitteleuropäischer Simuliidenarten gelten, obwohl sie Lücken aufweisen und die Um-weltfaktoren man gels spezifischer Erfahrun g nach allgemeinlimnologischen Gesichts-punkten ausgewählt worden sind. Offen bleibt vor allem, wie jeder einzelne Faktorzu gewichten ist und wieweit er das Weibchen bei der Wahl des EiablagepIatzesbeeinflusst.
Zur Frage der anthropogenen Einflüsse auf die Simuliidenfauna gehen die Mei-nungen der Autoren stark auseinander. RUBZOW (1959-1964) meldet eine Abnahmeder Kriebelmückendichte, die er auf die Eutrophierung der Flüsse zurückführt, ins-besondere durch die Zelluloseindustrie, ohne aber zu präzisieren, ob auf eine direktetoxische Wirkung chemischer Stoffe auf die Präimaginalstadien oder aber darauf, dassdie Eiablage- und Anheftungssubstrate durch exzessiven Algenbewuchs zugedecktwerden. WILHELM[ (1920) sieht in der Eutrophierung der Gewässer einen die Simu-liiden be günsti genden Faktor. THOMAS (1975) führt das reichliche Gedeihen sub-merser Makrophyten im Hochrhein, vor allem von Ranunculus fiuitans, auf die Zu-nahme eutrophierender Stoffe, in erster Linie Phosphat, zurück. Er meldet ebenfallseinen dichten Besatz dieser Pflanzen mit Larven und Puppen von Kriebelmücken,trotz der Toxizität des Wasserhahnenfusses für Wirbeltiere. Demnach mag die Dichtedes Makrophytenbewuchses die beiden Massenauftreten der Mücken in den Jahren1968 (EcKERT et al., 1969) und 1973 be günstigt haben. Daraus geht hervor, dassanthropogene Faktoren die Simuliidenfauna je nach der Art des Gewässers verschie-den beeinflussen können.
Nachdem in der ganzen Schweiz immer mehr Kläranlagen den Betrieb aufnehmen,
Jahrgang 123
R. GLATTHAAR. Verbreitung und Ökologie der Kriebelmiicken 119
dürften in naher Zukunft einige direkte und indirekte, bei der Komplexität lotischerÖkosysteme schwieri g vorhersehbare Änderun gen in der benthischen Fauna ein-treten, auch bei den Kriebelmiicken.
9. ZusammenfassuHg
1. Im Rahmen einer Querschnittsuntersuchun g schweizerischer Fliess gewässer undeiner turnusmässi gen Probenahme an ausgewählten Stellen im Kanton Zürich (Ta-belle 1) wurden in den Jahren 1973 und 1974 Daten von etwa 30 Umweltfaktorenerhoben (Tabelle 2), die auf die Präimaginalstadien und — in hier nicht untersuchtemMass — auf die Imagines einwirken. An denselben etwa 200 Probestellen wurden auchetwa 300000 Simuliiden eingesammelt, meist Präimaginalstadien, gele gentlich auchschwärmende Ima gines mit Hilfe eines Keschers.
2. Die aufgrund der vorliegenden Fänge erstellte Artenliste umfasst 28 Arten, vondenen 21 zum ersten Mal für die Schweiz nachgewiesen worden sind.
3. Die geo graphische Verbreitung dieser Arten in der Schweiz und die Fund-umstände werden beschrieben.
4. Die Phänologie von 13 häufigen Arten wird dar gestellt (Tabelle 3). Die Zahl derjährlichen Zyklen beträ gt 1 bis 4.
5. Es wird eine subjektive Einteilung der 17 häufi gsten Arten in fünf makroöko-logische Gruppen vorgeschlagen:
I. montane Gruppe, Bergbachbewohner,II. silvatische Gruppe, Waldbachbewohner,
IIl. potamale Gruppe, Bewohner grösserer Flachlandflüsse,IV. sublacustrische Gruppe, Bewohner von Seeausflüssen,V. campestrische Gruppe, Bewohner von Feld- und Wiesenbächen.
6. Die genannten fünf Gruppen werden mit den Befunden einer Chiqnadrat-Asso-ziationsanalyse, einer Cluster-Analyse mit den 350 er g iebi gsten Proben sowie mit einerCluster-Analyse mit allen Milieufaktoren verglichen. Sie werden von allen drei Ana-lysen zu verschiedenen Teilen bestäti g t (Tabelle 8).
7. Der Einfluss einzelner Milieufaktoren auf Verteilun g und Abundanz der 16 häu-fi gsten Arten wird anhand einer multiplen Regression Vund von Streudiagrammenerörtert.
Von diesen Analysen sowie von Einzelbeobachtungen an verschiedenen Probe-stellen kommen Hinweise, dass die physikalischen Faktoren stärkeren Einflnss aus-üben als die hydrochemischen.
Eine Cluster-Analyse der Zürcher Probestellen anhand der physikalischen und derhydrochemischen Faktoren unterstützt diesen Befund.
B. Die Fauna der-Thur zei g t im wesentlichen das aufgrund der Funde in anderenVoralpenflüssen postulierte Artenspektrum. Die Gliederun g der Thur wird mit jenerder Töss und der Fulda (Bundesrepublik Deutschland) verglichen.
Seeausflüsse wie die Glatt und der Hüttwilerseebach nei gen zu einer inversenSequenz. Die Zusammensetzung der Simuliidenfauna folgt weitgehend den physika-lischen Gegebenheiten.
120 Vierteljahrsschrift der Naturforschenden Gesellschaft in Zürich 1978
9. Die multiple Regression macht das Vorkommen verschiedener Simuliidenartenan Orten wahrscheinlich, wo sie in einigen Fällen bei späterer Suche tatsächlich gefun-den worden sind. Auch 17 in den Jahren 1973 und 1974 kriebelmiickenfreie Probe-stellen werden in Anbetracht späterer Funde durch die Cluster-Analyse wie auchdurch die multiple Regression gut charakterisiert.
10. Der Einfluss der menschlichen Zivilisation auf Verteilun g und Abundanz derSimuliiden wird diskutiert.
Résumé
1. Durant les annees 1973 et 1974 des echantillons ont ete pris et des dates ont ete levees concer-nant 30 mesures de l'envlronnement physique et chimique des eaux courantes (table 2) qui prennentinfluence sur les stades preimaginaux et les imagos n'etant pas comprises dans ce travail. L'activitescientifique s'etendait sur 200 emplacements suisses environ, dont 25 situes au canton de Zurich quifurent visites regulierement, les autres, repandus sur presque toute la Suisse, une fois ou a reprisessporadlques. Approximativement 300000 individus furent ramasses, surtout des stades preimaginaux,quelquefois des imagos essaimantes ä l'aide d'un filet ä papillons (table l).
2. La liste des especes etablie a base de ces prises contient 28 especes, dont 21 ont ete signaleespour la premiere fois en Suisse.
3. La repartition geographlque de ces especes en Suisse et les circonstances trouvees dans cesendroits observes sont decrites.
4. La phenologie des 13 especes frequentes est decrite (table 3). Le nombre des cycles annuelsest de un ä quatre.
5. Une division ä base d'impressions subjectives des 17 especes les plus frequentes est proposee,aboutissant a cinq groupes macro-ecologiques:
I. groupe montagnard-alpestre,II. groupe sylvatique, forestier,
III. groupe potamal, fluvial,IV. groupe sublacustre, des deversoirs,V. groupe champetre.
6. Les cinq groupes mentionnes ci-dessus sont compares aux resultats d'une analyse d'associationsA. base de Chi-carre, d'une analyse ä clusters contenant les 350 echantillons les plus efficaces ainsiqu'ä une analyse ä clusters a base de toutes les mesures des conditions environnementales. Le groupe-ment propose est confirme par les trois analyses dans differents aspects (table 8).
7. L'influence de composantes isolees du milieu sur la distribution et l'abondance des 16 especesles plus frequentes est discutee au moyen dune regression multiple et de diagrammes a dispersion.
Les analyses et divers observations isolees supportent l'hypothese que les composantes physiquesexercent plus d'influence que les conditions hydrochimiques.
Une analyse ä clusters portant sur les endroits zurichois ä base des facteurs physiques et hydro-chimiques confirme egalement cette hypothese.
8. La Thur a ete echantillonnee de la fontaine ä l'embouchure (Rhin). Son spectre simulidien seconforme aux associations proposees; il est compare à celui de la Fulda (Allemagne occidentale) etä celui de la Töss.
Les deversoirs de lacs, par exemple la Glatt et le Hüttwilerseebach, ont souvent une suite de lafaune renversee. La composition de la faune simulidienne se conforme ä grand mesure aux conditionsphysiques.
9. La regression multiple rend probable la presence de certaines especes a des endroits où ellesont ete trouvees dans plusieurs cas lors de recherches ulterieures. Dix-sept emplacements trouvesexempts de Simulies sont bien ranges par la regression multiple aussi bien que par l'analyse à clusters,vu quelques trouvailles ulterieures.
10. L'influence de la civilisation humaine exercee sur la dlstribution et l'abondance des Simuliesest discutee.
Jahrgang 123 R. GLATTHAAR. Verbreitung und Ökologie der Kriebelmücken 121
Summary
l. During the years 1973 and 1974 samples have been taken and data of 30 physical and hydro-chemicat factors of running waters have been recorded (table 2) concerning the influence upon thepreimaginal stages and (not investigated) upon the imagos of blackflies (Diptera, Simuliidae). About200 sampling places were chosen spread over nearly whole Switzerland (table l); 25 of them lnthe Zurich area were sampled regularly throughout the two years, the others once or sporadically.Approximatively 300000 individuals have been collected, especially preimaginat stages; on someoccasions swarming imagos were netted, too.
2. The list of species based upon these samples contains 28 species of which 21 are recorded forthe first time in Switzerland.
3. The geographic distribution of these species in Switzerland is given and the local conditions ofimportant sites are stated.
4. The phenology of 13 frequent species is presented (table 3). The number of annual cycles isbetween one and four.
5. A division of the 17 most frequent species based upon subjective impressions is proposed, lead-ing to five macro-ecological groups:
I. mountain group,II. silvatic group, forest group,
III. river group,IV. sublacustric group, living in outflows,V. country brook and stream group.
6. The five groups mentioned above are compared with the results of a Chi-square analysis ofassociations, of a cluster analysis including the 350 richest samples as well as with a cluster analysisincluding all environmental factors. The proposed grouping is confirmed in different parts by all thethree analyses (table 8).
7. The influence of isolated environmental factors upon the distribution and abundance of the16 most frequent species is discussed by means of a multiple regression and of scattering diagrams.
These analyses as wetl as isolated qualitative observations at some sampling sites hint at a majorrole of physical factors rather than of chemical ones.
A ctuster analysis including the sampling sites of the Zurich area basing upon the physical andhydrochemical factors confirms this result.
8. The Thur river has been sampled from its source to its estuary (Rhine river). Its fauna cor-responds mainly with the pattern yielded by the association analysis. It is compared with that ofthe Fulda river (Western Germany) and with that of the Töss stream.
Outflows of lakes such as the Glatt stream and the Hüttwilerseebach stream often show an inversesequence of the fauna, following mainly the physical conditions.
9. The multiple regIession gives cues of probable occurrence for some blackfly species at siteswhere they do have been found in further research work. Seventeen sites without blackflies during1973 and 1974 are well characterized by a cluster analysis as well as by the multiple regression, con-sidering later findings.
10. The influence of human civilization upon the distribution and abundance of the blackflies isdiscussed.
10. Literaturverzeichnis
BARANOV, N. (1939): Stand der Kolumbatscher-Mückenforschung in Jugoslawien. Z. Parasitenkde.11, 215-234.
CARLSSON, G. (1962): Studies on Scandinavian Blackflies. Opusc. Entomol., Suppl. 21, l-280.— (1967): Environmentat factors influencing blackfly populations. Bull. Wld. Health Org. 37, 139-
150.— (1968): Benthonic fauna of African watercourses with special reference to blackfly populations.
Scand. Inst. Afr. Studies Res. Rep. 3, l-13.
122
Vierteljahrsschrift der Naturforschenden Gesellschaft in Zürich 1978
CHUTTER, F. M. (1968): On the Ecology of the fauna on stones in the current in South African riverssupporting a very large Sinurlium population. Journ. appl. Ecol. 5, 531-561.
DAMES, D. M. (1961): Twdnnia Irvdroides NOVAK (Diptera, Simuliidae) from the Swiss Alps. Mittlg.Schweiz. Entomolog. Ges. 34, 269-270.
DAVIES, L. (1957): A new Prosimuliun: species from Britain, and a re-examination of P. hirtipes(FRIES) from the holarctic region. Proc. Royal Entomolog. Society London (B) 26, l-10.
— (1966): The Taxonomy of British blackflies (Diptera, Simuliidae). Transact. Royal Entomolog.Soc. London 118, 413-511.
DEMMERLE, S. D. (1966): Über die Verschmutzung des Rheines von Schaffhausen bis Kaiserstuhl.Vierteljahrsschr. Naturf. Ges. Zürich 111, 155-224.
DICE, L. R. (1945): Measures of Ecologic Associations between species. Ecology 26, 297-302.— (1948): Application of statistical methods to the analysis of ecologic association between species
of birds. Amer. Midland Naturalist 39, 174-178.DORIER, A. (1961): Sur la repartition des Simuliidae (Diptera) des Alpes francaises. Verh. Int. Verein.
Limnol. 14, 369-371.DORIER, A., P. GRENIER (1959): Une nouvelle Simulie (S. degrangei n. sp.) (Diptera, Simuliidae)
recoltee en Savoie. Tray. Lab. Hydr. Pisc. Grenoble 51, 195-204.ECKERr, J., H. GLOOR, E. KARBE, W. RUEHM (1969): Todesfälle durch Kriebelmücken (Diptera,
Simuliidae) bei Rindern in der Schweiz. Schweiz. Arch. Tierheilkde. 111, 447-455.EDWARDS, F. W. (1933): On three European species of Simalium. Ann. Naturhist. Mus. Wien 46,
255-256.Eidg. Departement des Innern, ed. 1974.. Vorläufi ge Empfehlungen über die regelmässige Unter-
suchung der schweizerischen Oberflächengewässer, 29. 5. 1974.ENIGK, K. (1955): Vorkommen und Bekämpfung der Kriebelmücken in Deutschland. Monatsh. wisst
prakt. Tierheilkde. 7, 241-253.GALLI-VALERIO, B. (1927): Beobachtun gen über Culiciden, nebst Bemerkungen über Tabaniden und
Simuliiden. Zentralbl. Bakteriol. (Abt. I, Orig.) 102, 224-226.GRENIER, P. (1949): Contribution a l'etude biologique des Simuliides de France. Physiologia comp.
cecol. 1, 165-330.— (1953): Simuliidae de France et d'Afrique du Nord. Encyclop. Entomol. Ser. A, 29, l-170.GRUNEWALD, J. (1972): Die hydrochemischen Lebensbedin gun gen der präimaginalen Stadien von
Boophthora ervthrocephala DEGEER (Diptera, Simuliidae). I. Freilanduntersuchungen. Z. Tropen-med. Parasitol. 23, 432-445.
— (1973): Die hydrochemischen Bedingungen ..., II. Die Entwicklung einer Zucht unter experimen-tellen Bedingungen. Z. Tropenmed. Parasitol. 24, 232-249.
HARNISCH, O. (1951): Hydrophysiologie der Tiere. Elemente zu ihrem Aufbau, in: Die Binnengewäs-ser, hg. A. THIENEMANN, Bd. 19, Stuttgart, Schweizerb.
ILLIES, J. (1953): Die Besiedlung der Fulda (insbesondere des Benthos der Salmonidenregion) nachdem jetzigen Stand der Untersuchung. Jber. Limnolog. Flussstat. Freudenthal, Göttingen, 5, 1-28.
— (1961): Versuch einer allgemeinen biozönotischen Gliederun g der Fliess gewässer. Internat. Revueges. Hydrobiol. 46, 205-213.
KNOZ, J. (1963): Poznámy k nekolika druhdm muchnickovitych (Diptera, Simuliidae), noch proüzemi Ceskoslovenska. Zool. Listy 12, 343-351.
— (1965): To identification of Czechoslovakian blackflies (Diptera, Simuliidae). Folia Fac. Sci. Nat.Univ. Purk. Brunensis 6, (5), l-54.
KUHN, H. (1952): Über das Vorkommen von Kriebelmückenlarven (Sin:ulium spec.) in Bächen aufdem Stadtgebiet Zürichs. Schweiz. Zschr. Hydrol. 14, 431 142.
MERKENS, J. C., K. M. DOWNING (1957): The effect of tension of dissolved oxygen on the toxicityof unionized ammonia to several species of fish. Ann. appl. Biol. 45, 521-527.
MOERGELI, W. (1974): Limnolo gische und morphologische Beziehun gen zwischen Wasser und Ge-rinnebett im Unterlauf der Thur. Vierteljahrsschr. Naturf. Ges. Zürich 119. 23-123.
MUEHLENBERG, M. (1976): Freilandökologie. Quelle & Meyer, Heidelberg, 214 pp.MUIRHEAD-THOMSON, R. C. (1970): The potentiatin g effect of pyrethrins and pyrethroids on the action
of organophosphorous larvicides in Simulium control. Transact. Royal Soc. Trop. Med. Hyg. 64,895-906.
Jahrgang 123 R. GLATTHAAR. Verbreitung und Ökologie der Kriebelmücken 123
PERRET, P. (1977): Zustand der schweizerischen Fliessgewässer in den Jahren 1974/75 (Projekt Mapos).Eidg. Amt f. Umweltschutz/EAWAG. 276 pp., 12 Karten.
PLEISCH, P. (1970): Die Herkunft eutrophierender Stoffe beim Pfäffiker- und Greifensee. Vierteljschr.Naturf. Ges. Zürich 115, 127-229.
RAI, H. (1970): The improvement of the sewage treatment of the city of Winterthur and its influenceon the river Töss. Vierteljschr. Naturf. Ges. Zürich 115, 1-100.
RIVOSECCHI, L. (1963): Contributo alla conoscenza dei Simulidi italiani, VII. Reperti negli Appenninied in Sicilia. Riv. Parassitol. 24, 119-142.
— (1967): I Simulidi degli Appennini. Parassitologia 9, 129-304.RrvosEccHI, L., L. LIPPARONI (1965): Contributo alla conoscenza dei Simulidi italiani, XI. Alcune
specie nuove per la fauna italiana. Riv. Parassitol. 26, 31-44.RIVOSECCHI, L., R. CARDINAL! (1975): Contributo alla conoscenza dei Simulidi italiani, XXIII. Nuovi
dati tassonomici. Riv. Parassitol. 36, 55-78.ROTHFELS, K. H. (1956): Black flies: Siblings, sex, and species grouping. J. Heredity 47, 113-121.RUBZOW, I. A. (1959-1964): Simuliidae (Melusinidae). In: Die Fliegen der Paläarktischen Region,
Bd. II1/4, ed. E. LINDNER. Stuttgart, Schweizerbart. 689 pp.— (1967): Simuliidae. In: Limnofauna Europaea, ed. J. ILLIES, pp. 340-345.RUEHM, W. (1967): Zur Verbreitung und Bedeutung der blutsaugenden Simuliiden im Aller-Leine-
Gebiet. Z. angew. Entomol. 59, 403-424.— (1968): Zur Autökologie einiger Simuliidenarten. Z. angew. Entomol. 61, 466-471.— (1969a): Der Nachweis von Kriebelmücken (Diptera, Simuliidae) an Weidetieren und Brutstät-
ten. Vet.-med. Nachrichten 1969, Heft l, 33-41.— (1969 b): Zur Populationsdynamik der Kriebelmücken, insbesondere von Boophthora erythro-
cephala DEGEER und des Odagmia-ornata-Komplexes. Z. angew. Entomol. 63, 212-227.— (1970 a): Zur Steuerung der Kriebelmückenschäden durch einige Umweltfaktoren. Z. angew.
Entomol. 65, 253-258.— (1970b): Zur Dispersion der Larvenstadien und des Puppenstadiums von Boophthora erythro-
cephala DEGEER (Simuliidae). Z. angew. Entomol. 66, 311-321.— (1970c): Zur Phänologie von Boophthora er ythrocephala DE GEER (Simuliidae, Diptera). Z. angew.
Zool. 57, 385-408.— (1971): Eiablage einiger Simuliidenarten. Z. angew. Parasitol. 12, 68-78.— (1972): Zur Populationsdynamik von Boophthora erythrocep/tala DEGEER. Z. angew. Entomol.
pp. 3-54.SNEATH, P. H. A., R. R. SOKAL (1973) : Numerical Taxonomy. Its principles and practice of numerical
classification. Freeman, San Francisco, 573 pp.SOKAL, R. R., P. H. A. SNEATH (1963): Principles of numerical taxonomy. Freeman, San Francisco,
359 pp.SUPPERER, R., E. KUTZER (1967): Beiträge zur Kriebelmücken-Fauna (Diptera, Simuliidae) Öster-
reichs. III. Die Fauna des Ybbstales. Verh. zool.-bot. Ges. Wien 107, 25-27.THOMAS, E. A. (1975): Zur Kenntnis der Toxizität des Flutenden Hahnenfusses (Ranunculus fluitarrs
LAM.). Vierteljschr. Naturf. Ges. Zürich 120, 275-281.Verzeichnis der Fischerei-Reviere und der Fischerei-Pächter des Kantons Zürich, hg. von der Finanz-
direktion des Kantons Zürich, 1970.WASER, E., W. HUSMANN, G. BLOECHLIGER (1934): Die Glatt. Berichte Schweiz. Bot. Ges. 43,253-
388.WASER, E., G. BLOECHLIGER, E. A. THOMAS (1943): Untersuchungen am Rhein von Schaffhausen bis
Kaiserstuhl. Schweiz. Z. Hydrol. 9, 225-309.WASER, E., E. A. THOI.L4S (1944): Untersuchungen an der Thur 1940/41. Schweiz. Z. Hydrol. 10, 1-89.WILHELMI, J. (1920): Die Kriebelmückenplage. G. Fischer Verlag, Jena. 246 pp.ZIVKOVITCH, V. (1961): Rasprostranenost i znacaj simulida (Diptera, Simuliidae) u Jugoslaviji. Glas.
Sr. Acad. nauka i umes. 248 (odel. med. nauka) 16, 133-142.ZOBRIST, J., J. S. DAVis, H. R. HEGt (1976): Charakterisierung des chemischen Zustandes des Flusses
Glatt. Gas, Wasser, Abwasser 56, 89-119.
124 Vierteljahrsschrlft der Naturforschenden Gesellschaft in Zürich 1978
ZWICK, H., W. RUEHM (1972): Erstnachweis von Simuliunz sublacustre DAVIES 1966 in Mitteleuropa.Z. angew. Entomol. 72, 429-434.
ZWICK, H. (1974): Faunistisch-ökolo gische und taxonomische Untersuchungen an Simuliidae (Dip-tera) mit besonderer Berücksichtigung der Arten des Fulda-Gebietes. Abb. Senckenberg. Naturf.Ges. 533, l-116.
— (1976): Zur Kenntnis der Kriebelmücken-Fauna (Simuliidae, Diptera) Österreichs. Z. Arbeits-gem. Österr. Entomolo gen 28, 73-77.
ZwicK, P. (1969): Beitrag zur Kenntnis der Plecopterenfauna der Fulda und ihres Einzugsgebietes inder Rhön und im Vogelsber g. Beitr. Naturkde. Osthessens, Fulda, 1, 65-76.