Page 1
Aus der Klinik und Poliklinik für Chirurgie
(Direktor: Univ.- Prof. Dr. med. Claus-Dieter Heidecke)
der Medizinischen Fakultät der Ernst-Moritz-Arndt-Universität Greifswald
Das Verhalten und der Einfluss von Thrombozyten und CCR4 in der
polymikrobiellen Sepsis
Inaugural – Dissertation
zur
Erlangung des akademischen
Grades
Doktor der Medizin
(Dr. med.)
der
Medizinischen Fakultät
der
Ernst-Moritz-Arndt-Universität
Greifswald
2010
vorgelegt von:
Appelt, Manja
geb. am: 08.12.1986
in: Schwedt/Oder
Page 2
Dekan: Prof. Dr. med. dent. Reiner Biffar
1. Gutachter: Prof. Dr. med. Stefan Maier
2. Gutachter: Prof. Dr. med. A. Greinacher
3. Gutachter: Prof. Dr. Bröker
4. Gutachter: Priv.-Doz. Dr. med. Nico Schäfer
5. Gutachter: Priv.-Doz. Kollmar
Ort, Raum: Seminarraum der Klinik für Thoraxchirurgie
Tag der Disputation: 28.10.2013
Page 4
IV
Inhaltsverzeichnis
Abkürzungsverzeichnis .................................................................................. VII
1 Einleitung ................................................................................................... 1
1.1 Thrombozyten................................................................................................ 2
1.1.1 Aufbau des Thrombozyten ...................................................................... 2
1.1.2 Aktivierung des Thrombozyten ................................................................ 5
1.1.3 Funktion des Thrombozyten.................................................................... 7
1.1.4 Thrombozyten in der Sepsis ................................................................... 8
1.2 Pathogenese der Sepsis ............................................................................... 9
1.3 Sepsismodelle ............................................................................................. 12
1.3.1 Exogene Verabreichung von Toxinen und Bakterien ............................ 12
1.3.2 Cecal Ligation and Puncture (CLP) ....................................................... 13
1.3.3 Colon Ascendens Stent Peritonitis (CASP) ........................................... 13
2 Fragestellung der Arbeit ........................................................................... 15
3 Material und Methoden ............................................................................. 16
3.1 Material ........................................................................................................ 16
3.1.1 Reagenzien und Chemikalien ............................................................... 16
3.1.2 Kits ........................................................................................................ 18
3.1.3 Primer ................................................................................................... 18
3.1.4 Antikörper.............................................................................................. 18
3.1.5 Puffer und Lösungen ............................................................................. 19
3.1.6 Laborgeräte ........................................................................................... 20
3.1.7 Verbrauchsmaterialien .......................................................................... 21
3.2 Methodik in vivo ........................................................................................... 22
3.2.1 Genehmigung der Tierversuche ............................................................ 22
3.2.2 Tiere und Tierhaltung ............................................................................ 22
3.2.3 Tiermodelle ........................................................................................... 23
Page 5
V
3.2.3.1 Anästhesie ...................................................................................... 23
3.2.3.2 Colon Ascendens Stent Peritonitis (CASP) .................................... 23
3.2.3.3 Schmerztherapie ............................................................................ 25
3.2.4 Organentnahme .................................................................................... 25
3.3 Zellbiologische Methoden mit humanen Zellen ........................................... 26
3.3.1 Abnahme und Präparation der Blutproben ............................................ 26
3.3.2 Stimulation von Vollblut ......................................................................... 26
3.3.3 Stimulation von plättchenreichem Plasma (PRP) .................................. 26
3.4 Zellbiologische Methoden mit murinen Zellen ............................................. 27
3.4.1 Isolation muriner Thrombozyten ............................................................ 27
3.4.2 Aufarbeitung muriner Lymphozyten ...................................................... 27
3.4.2.1 Isolation von PBMCs aus dem peripheren Blut .............................. 27
3.4.2.2 Isolation von CD4+-Zellen aus der Milz........................................... 28
3.5 Molekularbiologische Methoden .................................................................. 29
3.5.1 RNA-Isolation ........................................................................................ 29
3.5.2 Reverse Transkription und Polymerasekettenreaktion .......................... 31
3.6 Proteinbiochemische Methoden .................................................................. 32
3.6.1 Durchflusszytometrie humaner Zellen ................................................... 32
3.6.1.1 Vollblutfärbung für die Durchflusszytometrie .................................. 33
3.6.1.2 Färbung des PRP ........................................................................... 34
3.6.2 Durchflusszytometrie muriner Zellen ..................................................... 34
3.6.3 Fibrinogen-ELISA .................................................................................. 35
3.6.4 Immunhistologie .................................................................................... 35
3.6.4.1 Anfertigen von histologischen Präparaten ...................................... 35
3.6.4.2 Färbungen ...................................................................................... 35
3.7 Statistische Auswertung .............................................................................. 38
4 Ergebnisse ............................................................................................... 39
4.1 In vitro Versuche mit humanen Zellen ......................................................... 39
Page 6
VI
4.1.1 Stimulation von plättchenreichem Plasma ............................................ 39
4.1.1.1 Stimulation mit TRAP-6 .................................................................. 40
4.1.1.2 Stimulation mit TRAP-6 und LPS ................................................... 41
4.1.2 Stimulation von Vollblut ......................................................................... 44
4.1.2.1 Granulozyten .................................................................................. 46
4.1.2.2 Monozyten ...................................................................................... 49
4.2 Versuche mit murinen Thrombozyten .......................................................... 52
4.2.1 Thrombozytenzahl nach CASP ............................................................. 52
4.2.2 Immunhistologischer Vergleich septischer und unbehandelter Organe 52
4.2.2.1 Ergebnisse von Lunge und Milz in CCR4-/- und C57BL/6 ............... 53
4.2.2.2 Ergebnisse von Leber..................................................................... 56
4.2.2.3 Ergebnisse der Leber mit CD42c , CD62P und Gr-1 ...................... 57
4.2.3 Fibrinogen-ELISA .................................................................................. 60
4.3 CCR4-Regulation nach CASP ..................................................................... 62
4.3.1 RT-qPCR muriner PBMCs .................................................................... 62
4.3.2 RT-qPCR muriner CD4+-Zellen ............................................................ 65
5 Diskussion ................................................................................................ 67
5.1 In vitro Versuche mit humanen Zellen ......................................................... 67
5.2 In vivo Versuche nach CASP ...................................................................... 70
5.3 CCR4-Regulation nach CASP ..................................................................... 74
6 Zusammenfassung ................................................................................... 77
Literaturverzeichnis ........................................................................................ 79
Verzeichnis der Abbildungen und Tabellen .................................................... 86
Eidesstattliche Erklärung ................................................................................ 88
Lebenslauf ..................................................................................................... 89
Danksagung ................................................................................................... 91
Page 7
VII
Abkürzungsverzeichnis
ACDA Acid Citrate Dextrose solution A
AK Antikörper
BSA Bovines Serumalbumin
CARS compensatory anti-inflammatory
response syndrome
CASP Colon Ascendens Stent Peritonitis
CCL17 Thymus- and activation-regulated
chemokine
CCL22 Macrophage-derived chemokine
CCR4 Chemokinrezeptor 4
C57BL/6 C57 Black 6 – Maus
CLP Cecal ligation and puncture
DAMP Damage associated molecular
pattern molecules
DIC Disseminierte intravasale Gerinnung
EDTA Ethylendiamintetraessigsäure
FACS Fluorescence-activated-cell-sorter
FITC Fluoresceinisothiocyanat
FCS Fetales Kälberserum
GAPDH Glycerinaldehyd-3-phosphat-
Dehydrogenase
GP Glykoprotein
GPIb/V/IX Rezeptor für vWF auf Thrombozyten
GPIIb/IIIa Fibrinogenrezeptor auf Thrombozyten
16G 16 Gauge
HMGB1 High-Mobility-Group-Protein B1
IL Interleukin
IFN Interferon
i.p. intraperitoneal
LPS Lipopolysaccharid
MDC Macrophage - derived chemokine
Page 8
VIII
MODS Multi organ dysfunction syndrome
NETs Neutrophil extracellular traps
NFκB nuclear factor of activated B-cells
OCS open canalicular system
PAC-1 Antikörper gegen aktiviertes GPIIb/IIIa
PAMP Pathogen - associated molecular
patterns
PAR 1-6 Protease-activated Rezeptor 1-6
PCR Polymerasekettenreaktion
PBS Phosphate buffered Saline
PBMC Peripheral Blood Mononuclear Cell
PDGF Platelet derived growth factor
PE Phycoerythrin
PF-4 Platelet factor 4
POD Peroxidase
PRP Plättchenreiches Plasma
PRR Pattern recognition receptor
PSGL-1 P-Selektin Glykoprotein Ligand-1
RGDS Arg-Gly-Asp-Ser
RT Raumtemperatur
TARC Thymus- and activation-regulated
chemokine
TF Tissue Factor
TGF Transforming growth factor
TLR Toll-like Rezeptor
TNF Tumornekrosefaktor
TRAP-6 Thrombin receptor agonist peptide-6
vWF Von-Willebrand-Faktor
WT Wildtyp
Page 9
Kapitel 1: Einleitung
1
1 Einleitung
Trotz intensiver Forschung und zahlreichen therapeutischen Möglichkeiten
stellt das Krankheitsbild der Sepsis noch immer ein großes Problem auf
intensivmedizinischen und chirurgischen Stationen dar. Die Mortalität ist sehr
hoch und die weltweite Inzidenz steigend [1-3]. Allein in Deutschland
erkranken jährlich 76.000 Patienten an Sepsis und septischem Schock, damit
liegt die Prävalenz auf deutschen Intensivstationen bei 12,4% für Sepsis und
11% für schwere Sepsis [3, 4]. Die Letalität der schweren Sepsis beträgt trotz
stationärer Behandlung und Medikation 55,2%. Es sterben pro Jahr etwa
60.000 Patienten an einer Sepsis. Damit ist sie hinter der koronaren
Herzkrankheit und dem Myokardinfarkt die dritthäufigste Todesursache [4].
Auch wenn in den letzten 20 Jahren das Verständnis pathophysiologischer
Zusammenhänge auf dem Gebiet der Sepsis stark erweitert wurde, stehen
Kliniker noch immer vor den Problemen der rechtzeitigen Diagnose und
entsprechenden Behandlung der Sepsis. Besonders bei den frühen Stadien
der Sepsis ist es wichtig entsprechend einzugreifen, doch gerade diese
werden diagnostisch häufig zu spät erkannt [5].
Definitionsgemäß wird die Sepsis heute vereinfacht als generalisierte
hyperinflammatorische Reaktion auf eine Infektion bezeichnet, die mit SIRS
(systemic inflammatory response syndrome) abgekürzt wird [6]. Dabei sind
Bakterien neben Viren und Pilzen die Hauptauslöser der Infektion und führen
zur Aktivierung von Rezeptoren der angeborenen Immunität. Durch Verlust
von Kontrollmechanismen führt die ausgedehnte Immunantwort schließlich zur
Schädigung des eigenen Organismus. Allerdings überwiegt nur initial diese
proinflammatorische Phase, zusätzlich kommt es in den späten Stadien der
Sepsis gegenregulatorisch zur Antiinflammation, die man als CARS
(compensatory anti-inflammatory response syndrome) bezeichnet, bis hin zur
Immunparalyse. Diese gegensätzlichen Erscheinungen während der Sepsis
und das teilweise Überlappen der beiden Phasen erschweren die Medikation
[7]. Neben dem Ungleichgewicht des Immunsystems gerät auch die
Hämostase außer Kontrolle. Auf der einen Seite kommt es zur massiven
Aktivierung der Gerinnung, die bis hin zum Vollbild der disseminierten
intravasalen Gerinnung (DIC) führen kann. Auf der anderen Seite kommt es
Page 10
Kapitel 1: Einleitung
2
zur Aktivierung von Thrombozyten und zur Thrombozytopenie [8]. Sowohl die
gestörte Hämostase, als auch der Abfall von Thrombozyten werden deshalb
unter anderem als diagnostische Marker für die schwere Sepsis verwendet.
Bedeutend sind diese oft vernachlässigten Aspekte, weil eine schwere DIC mit
erhöhter Thrombenbildung und Verbrauchskoagulopathie zur eingeschränkten
Mikrozirkulation der Organe und damit zu deren Versagen führen kann. Die
Thrombozytopenie und der Verbrauch von Gerinnungsfaktoren führen
außerdem zur Hypokoagulabilität und so zur Blutungsgefahr für den Patienten.
Da der Thrombozyt im Krankheitsbild Sepsis selten in Betracht gezogen wird,
obwohl seit langem bekannt ist, dass er mit dem Immunsystem interagiert, soll
seine Funktion in dieser Arbeit näher betrachtet werden.
1.1 Thrombozyten
1.1.1 Aufbau des Thrombozyten
Thrombozyten sind mit einem Durchmesser von 2-5 µm und einem Volumen
von 6-10 fl die kleinsten Zellen im menschlichen Organismus. Die Größe wird
aber mit der immensen Plättchenanzahl von 150000 bis 300000 pro Mirkoliter
Blut ausgeglichen. Diese Menge zirkulierender Thrombozyten entspricht etwa
dem 50fachen an Leukozyten.
Sie haben eine Lebensdauer von etwa 7-10 Tagen im Blut und werden dann
von der Leber oder Milz abgebaut. Sie gelangen in die Zirkulation als kernlose,
diskoide Abkömmlinge der Megakaryozyten, mehrkernigen Riesenzellen im
Knochenmark. Thrombozyten besitzen zwar keinen Zellkern, aber sonst
beinhalten sie durch die Abschnürung vom Megakaryozyt alle
überlebenswichtigen Organellen, sowie zahlreiche Rezeptoren und residuelle
mRNA mit allen notwendigen Mechanismen zur Neusynthese von Proteinen
[9-12].
Der Aufbau der diskoiden Zelle ist sehr homogen, aber dennoch recht
komplex. Die Plasmamembran weist viele Fältelungen auf zur
Oberflächenvergrößerung und dringt in das Innere des Thrombozyts ein zur
Bildung seines einzigartigen kanalikulären Systems (OCS, open canalicular
system). Dieses OCS zieht sich schwammartig durch die Zelle und dient zur
zusätzlichen Speicherung von Proteinen der Glykokalix. Außerdem ist es
beteiligt an Endozytosevorgängen und bei der Sekretion von Proteinen aus
Page 11
Kapitel 1: Einleitung
3
den Granula. Neben dem offenen kanalikulären System, findet man in den
kleinen Zellen auch ein geschlossenes kanalikuläres System, das vom rauen
endoplasmatischen Retikulum des Megakaryozyts abstammt. Es beinhaltet
Calcium und zahlreiche Enzyme und übernimmt damit die Kontrolle über
Aktivierungsvorgänge des Thrombozyts und verschiedene metabolische
Funktionen wie die Thromboxansynthese [13]. Weiterhin enthält der
Thrombozyt Speichergranula, die α-, δ- und λ-Granula, die verschiedene
Aktivierungsmarker und Botenstoffe enthalten und nach Aktivierung über
Exozytose freigesetzt werden [14]. Während die λ-Granula Lysosomen
darstellen, enthalten die dichten δ-Granula vor allem niedermolekulare
Substanzen wie Calcium, Nukleotide und Serotonin. Die α-Granula speichern
vorwiegend Proteine wie Plättchen-Faktor 4 (PF4), P-Selektin (CD62P),
Matrixproteine wie Fibrinogen und Fibronektin sowie Wachstumsfaktoren wie
PDGF (platelet derived growth factor) und TGF-β (transforming growth factor).
Inhalte der Granula diffundieren nach Exozytose entweder in den
Extrazellulärraum oder werden als Rezeptoren auf der
Thrombozytenoberfläche exprimiert.
Abb. 1: Schematische Darstellung eines Thrombozyts mit all seinen Organellen
Page 12
Kapitel 1: Einleitung
4
Zu sehen sind neben dem Membransystem, einmal dem offenen kanalikulären System (OCS)
und dem geschlossenen kanalikulären System (DTS), die Mitochondrien und die drei
spezifischen Granula (Bildquelle: Rendu et. al) [13].
Die δ-Granula sind kleiner und dichter als die stärker granulierten, irregulären
α-Granula. Die Lysosomen (λ-Granula) weisen eine mittlere Größe und
komplette dichte Füllung auf [13, 15].
Neben dem generellen Aufbau ist vor allem die Expression zahlreicher
Oberflächenmoleküle für die Funktion und Interaktion des Thrombozyten von
Bedeutung. Zu finden sind konstitutiv exprimierte, aber auch induzierbare
Rezeptoren. In den letzten Jahren zeigte sich, dass Thrombozyten nicht nur
funktionelle Rezeptoren für ihre Rolle in der Hämostase aufweisen, sondern
auch viele Oberflächenproteine, die für eine Funktion im Immunsystem
sprechen.
In der Hämostase sind vor allem die Adhäsionsmoleküle von Bedeutung,
spezifische adhäsive Glykoproteine (GP). Die wichtigsten sind hier der
Rezeptor GPIb, der durch Multimere des von Willebrand Faktor (vWF) zum
GPIb/V/IX-Komplex konformiert und so den vWF unter hoher Schubspannung
binden kann. Dieser Vorgang ist wichtig für die initiale Adhäsion der
Thrombozyten an die subendotheliale Matrix, die bei Gefäßschäden freiliegt.
Der vWF, der unter anderem von Endothelzellen der Gefäßintima gebildet
wird, dient dabei als Bindeglied zwischen Thrombozyten und dem bei
Gefäßschäden freiliegendem Kollagen. Bedeutend für die Interaktion zwischen
Thrombozyten ist der Rezeptor für Fibrinogen. Dieser Rezeptor mit der
Bezeichnung GPIIb/IIIa (αIIbβ3) kann erst nach Plättchenaktivierung und
folgender Strukturänderung Fibrinogen binden und ist dann zuständig für die
Thrombusbildung. Pac-1, ein spezifischer Antikörper, der nur an die aktivierte
Form des GPIIb/IIIa-Rezeptors bindet, wird als spezifischer
Aktivierungsmarker bei Thrombozyten genutzt.
Neben den Glykoproteinen besitzen Thrombozyten weitere Rezeptoren, die
eine Rolle im Immunsystem nahe legen. Sie exprimieren pattern recognition
receptors (PRR) wie toll-like Rezeptoren [16, 17] und exozytieren bei
Aktivierung durch mikrobielle Strukturen Mediatoren wie Chemokine und
Zytokine. Als wichtigste Beispiele sollten hier Chemokine wie RANTES
(regulated upon activation, normal T cell expressed and secreted), MIP-1α
Page 13
Kapitel 1: Einleitung
5
(macrophage inflammatory protein 1α) , PF4 und das Zytokin Il1-β genannt
werden. Neben der Ausschüttung von Botenstoffen werden nach Aktivierung
Oberflächenproteine exprimiert, die eine Interaktion mit anderen Immunzellen
vermitteln. Eines der wichtigsten ist hier Adhäsionsmolekül und
Aktivierungsmarker P-Selektin, das nach Plättchenaktivierung aus den
α-Granula entlassen und auf die Membranoberfläche transportiert wird. An
P-Selektin adhärieren Leukozyten über PSGL-1 (P-Selektin Glykoprotein
Ligand-1) und können so auf Plättchenthromben entlang rollen. Der
Aktivierungsmarker wird nach Stimulation mit Thrombin vermehrt
ausgeschüttet. Diese Tatsache wird genutzt um die Funktion von
Thrombozyten zu überprüfen. Verwendet wird dazu auch das synthetische
Thrombinanalogon TRAP-6 (Thrombin receptor agonist peptide-6), das wie
Thrombin über den protease-activated receptor 1 (PAR-1) in sehr geringen
und über PAR-4 in höheren Konzentrationen stark aktivierend wirkt [18].
Ein weiteres Bindungsmolekül für Leukozyten ist CD40L, das auch aus den
α-Granula freigesetzt wird nach Stimulation der Plättchen mit typischen
Thrombozytenagonisten wie Thrombin, Kollagen oder ADP [19].
1.1.2 Aktivierung des Thrombozyten
Thrombozyten lassen sich über verschiedene Wege aktivieren.
Klassischerweise ist die Adhäsion zur subendothelialen Matrix der initiale
Schritt zur Plättchenaktivierung. Bei Beschädigung der Gefäßinnenwand wird
das darunterliegende Subendothel mit seinen Struktur- und
Adhäsionsproteinen wie Kollagen und von-Willebrand-Faktor (vWF) frei. Der
Thrombozyt adhäriert mit den entsprechenden oben genannten spezifischen
Oberflächenproteinen und wird dadurch aktiviert. Zusätzlich kann auch die
Bindung von spezifischen Agonisten wie Thrombin, Kollagen oder ADP an
thrombozytäre Rezeptoren zur starken Aktivierung führen, wobei Thrombin der
stärkste Thrombozytenaktivator überhaupt ist [20]. Nach der Aktivierung
kommt es zu einer Formänderung („shape change“) der Thrombozyten (Abb.
2), die elektronenmikroskopisch sichtbar wird in einer immensen
Oberflächenvergrößerung und Ausbildung von Pseudopodien. Außerdem
fusionieren die Granula mit der Plasmamembran und sezernieren ihren Inhalt
(Degranulation). Es kommt zur Ausschüttung von Sekretionsprodukten wie
Page 14
Kapitel 1: Einleitung
6
ADP, Thrombin oder Serotonin und zur Expression von Aktivierungsmarkern
wie P-Selektin. Die Aktivierung ist assoziiert mit einer Bindung von Fibrinogen
an den aktivierten GPIIb/IIIa, die eine Brückenbildung zwischen den
Thrombozyten vermittelt und letztendlich zur Aggregation führt.
Diese Thrombusbildung durch Thrombozyten nennt man auch primäre
Hämostase, während die anschließende Initiation der Gerinnungskaskade als
sekundäre Hämostase bezeichnet wird. Sie wird getriggert durch die
Degranulation und Ausschüttung von Thrombin aus den Thrombozyten. Die
Gerinnung endet in der Synthese von Fibrin und führt zu festen Fibrin-
Plättchen-Thromben. Die granulären Inhaltsstoffe lösen nicht nur die
Gerinnungskaskade aus, sondern führen auch zur Rekrutierung weiterer
Thrombozyten und deren Aktivierung, so dass es zur Amplifikation der
Prozesse kommt.
Abb. 2: Änderung der Thrombozytenform nach Aktivierung
Ruhende Plättchen weisen eine diskoide, homogene Form auf (A). Nach Aktivierung kommt
es zur Kontraktion und Umordnung des Zytoskeletts, resultierend in einer Ausbildung von
Pseudopodien und einer Formänderung (Bildquelle: Mony M. Frojmovic, John G. Milton [21]).
Die sekundäre Hämostase, die über die Gerinnungsfaktoren vermittelt wird,
wird neben Thrombozyten durch den tissue factor (TF) generiert. TF wird
normalerweise konstitutiv von subendothelialen Zellen exprimiert, die keinen
Kontakt zum Blut und damit zu den Gerinnungsfaktoren haben. Bei Verletzung
der Gefäßwand kommt der TF in Kontakt mit dem Blut und es kommt zur
Bindung von Faktor VII und damit zum Anstoß der Gerinnungskaskade. Außer
von den subendothelialen Zellen kann TF von im Blut zirkulierenden
Monozyten und Makrophagen nach deren Stimulation generiert werden.
Page 15
Kapitel 1: Einleitung
7
Stimulanzien, die dazu in der Lage sind, sind sowohl Zytokine wie IL-1 oder
TNF, als auch Lipopolysaccharid (LPS) nach Langzeitstimulation. Da dies eine
TF-abhängige Gerinnung ohne Gefäßverletzung bedeuten würde, spielt dieser
Prozess für Hämostasestörungen in der Sepsis eine besondere Rolle.
Thrombozyten tragen durch direkte adhäsive Interaktionen zu der
monozytären TF-Aktivität bei [22, 23].
Die kaskadenartige Aktivierung der Gerinnungsfaktoren endet in der Synthese
von Thrombin, was letztendlich von Fibrinogen Fibrin abspaltet. Fibrinogen ist
ein in der Leber gebildetes Protein, das neben seiner Funktion in der
Hämostase auch als Akute-Phase-Protein fungiert. Da es als Protein einfach
per ELISA im Plasma zu messen ist, wird es als Marker für verschiedene
Erkrankungen genutzt. Ein Absinken würde entweder für eine
Leberfunktionsstörung oder für einen sehr starken Verbrauch von Fibrinogen
sprechen, während ein Anstieg des Akute-Phase-Proteins bei
Entzündungsprozessen und nach operativen Traumen zu verzeichnen wäre.
Zu einem Verbrauch käme es beispielsweise bei starker Aktivierung der
Gerinnungskaskade, wie es bei disseminierter intravasaler Gerinnung der Fall
wäre. Aus diesem Grund zählt ein verminderter Fibrinogengehalt im Plasma
(<100 mg/dl) neben einer verringerten Thrombozytenzahl, einer Verlängerung
der Prothombinzeit und einer Erhöhung von Fibrinmarkern laut der
Internationalen Gesellschaft für Thrombose und Hämostase (ISTH) zu den
Nachweisparametern einer DIC [24].
1.1.3 Funktion des Thrombozyten
Wie aus der Struktur des Thrombozyten bereits hervorgeht, besteht seine
Hauptfunktion in der Aufrechterhaltung der Hämostase, also der Stillung von
Blutungen. Durch seine einzigartige Struktur ist er bestens dafür geeignet
kleine Gefäßverletzungen abzudichten, indem er über Fibrinogen
Querbrücken und Bindungen untereinander ausbildet. Die primäre
Defektheilung durch den Thrombozyt wird unterstützt durch die Aktivierung
des Gerinnungssystem und der Stabilisierung des Thrombus durch Fibrin. Es
kommt zur komplexen Verdichtung durch die Interaktion mit dem
Gerinnungssystem.
Page 16
Kapitel 1: Einleitung
8
Neben seine Rolle in der Hämostase wird seit einigen Jahren auch seine
Mitwirkung im Immunsystem diskutiert. Über entsprechende Rezeptoren
interagiert er sowohl mit den bakteriellen Pathogenen als auch mit den
rekrutierten Immunzellen. Thrombozyten werden im Entzündungsgeschehen
aktiviert und kommunizieren über Rezeptoren und die Ausschüttung von oben
genannten Mediatoren mit dem Immunsystem.
1.1.4 Thrombozyten in der Sepsis
Da Thrombozyten beteiligt sind an inflammatorischen Prozessen, ist es nahe
liegend, dass sie auch in der Sepsis einen Einfluss haben. Bekannte
Phänomene für das schwere Krankheitsbild sind die Thrombozytopenie und
die disseminierte intravasale Gerinnung (DIC).
Als Thrombozytopenie bezeichnet man einen Abfall der Blutplättchen auf
weniger als 150 x 109 Plättchen pro Liter Blut. Von diesem Thrombozytenabfall
sind etwa 35-44% der septischen Patienten betroffen. 12-15% weisen sogar
ein Absinken auf nur noch 50 x 109/l Zellen auf. Die septischen Patienten
erfahren diesen Zellrückgang innerhalb der ersten vier Tage auf der
Intensivstation. Das Ausmaß der Thrombozytopenie korreliert direkt mit der
Schwere der Sepsis [25]. Die Ursache für dieses Phänomen in der Sepsis ist
noch nicht vollständig geklärt.
Die systemische Inflammation führt außerdem zur Aktivierung der
plasmatischen Gerinnung und zur Hemmung der Fibrinolyse [26]. Durch die
Bildung von Fibrinablagerungen können eindringende Mikroorganismen und
die inflammatorische Antwort eingedämmt werden [27]. Eine überschießende
Gerinnung kann dagegen mikrovaskuläre Thromben und Organdysfunktion
nach sich ziehen, ein Krankheitsbild, das man dann als disseminierte
intravasale Gerinnung bezeichnet. Hauptaktivator der Gerinnung in der Sepsis
ist der in Makrophagen induzierbare Gewebefaktor, der sogenannte tissue
factor (TF). In der Sepsis anflutende bakterielle Strukturen wie LPS, aber auch
Zytokine wie TNF-α oder P-Selektin von aktivierten Thrombozyten führen zur
Generierung von TF.
Die folgende Verminderung von Gerinnungsfaktoren bei septischen Patienten
bezeichnet man auch als Verbrauchskoagulopathie und führt zu einer
verstärkten Blutungsneigung. Neben dem erhöhten Verbrauch kommt es auch
Page 17
Kapitel 1: Einleitung
9
durch verminderte Synthese und Verlust zum Absinken der
Gerinnungsfaktoren [28].
1.2 Pathogenese der Sepsis
Der klassische Begriff der Sepsis wurde bereits 1914 von Hugo Schottmüller
als hämatogene Generalisation einer Lokalinfektion geprägt. Ursache sind
hierbei vor allem die eindringenden Mikroorganismen. Erst Jahrzehnte später
führte eine Theorie von Lewis Thomas zum Weiterdenken im
pathophysiologischen Konzept der Sepsis. Sie besagte, dass nicht nur die
eindringenden Mikroorganismen an sich, sondern vor allem die
Abwehrreaktionen des Organismus für die Schädigung im Rahmen einer
Sepsis verantwortlich seien. Seine Theorie wurde schließlich bestätigt durch
eine Reihe von Studien und mündete in der Definition des systemic
inflammatory response syndrome (SIRS) durch Roger Bone [7, 29]. Am
Anfang dieser hyperaktiven, unkontrollierten Immunreaktion steht vor allem
das angeborene Immunsystem. Es reagiert auf exogene Schädigungen, die
neben Viren und Pilzen vor allem durch Bakterien verursacht werden. Das
Keimspektrum der Sepsis ist abhängig vom Ausgangspunkt der Infektion und
von verschiedenen Wirtsfaktoren wie beispielsweise einer bestehenden
Immunschwäche des Patienten. Trotzdem findet man generell bestimmte
Keime häufiger als andere. Die wichtigsten Vertreter sind die gram-negativen
natürlichen Darmbakterien wie E.coli, Klebsiellen und Enterokokken, sowie
gram-positive Staphylokokken und Streptokokken. Zellen des angeborenen
Immunsystems, wie Makrophagen, neutrophile Granulozyten und dendritische
Zellen können typische molekulare Muster auf diesen Mikroben erkennen,
binden und präsentieren. Diese konservierten Muster, die man auf den
Pathogenen findet, werden als PAMPs (Pathogen-associated molecular
pattern) bezeichnet. Außerdem werden bei Gewebsschäden vom Organismus
sogenannte Alarmine ausgeschüttet, deren bekanntester Vertreter wohl das
HMGB1 (High-Mobility-Group-Protein B1) ist. Alarmine werden nur von
nekrotischen Zellen und Immunzellen freigesetzt. PAMPs und Alarmine fasst
man unter dem Begriff DAMPs (Damage associated molecular pattern
molecules) zusammen. Sowohl die körperfremden Muster als auch die
eigenen Alarmstoffe werden von den Zellen über PRRs wie den toll-like
Page 18
Kapitel 1: Einleitung
10
Rezeptoren (TLRs) erkannt und gebunden. Bereits durch Antikörper oder
Komplementfaktoren markierte Bakterien werden durch Fc-Rezeptoren auf
Makrophagen und neutrophilen Granulozyten gebunden, phagozytiert und
durch Fusion mit Lysosomen beseitigt. Die TLR-ausgelöste Signalkaskade
resultiert in der Dislokation des Transkriptionsfaktors NFκB in den Zellkern und
führt zur Aktivierung hunderter verschiedener Gene, die für Proteine
proinflammatorischer Reaktionen kodieren. Neben NFκB werden auch noch
zahlreiche weitere Transkriptionsfaktoren zeitabhängig aktiviert [7].
Es kommt zur Synthese von Zytokinen und Chemokinen. In gesunden
Menschen werden nur sehr geringe Plasmakonzentrationen (pg/ml) detektiert,
während es in der Sepsis zum exzessiven Anstieg der Menge kommt.
Zytokinrezeptoren werden von fast allen Zellen exprimiert und finden sich
zusätzlich in löslicher Form im Plasma. Die wichtigsten Zytokine, die in der
Anfangsphase der Immunreaktion von Makrophagen ausgeschüttet werden,
sind TNF-α und IL-1. Die beiden Zytokine induzieren Leukozyten,
Endothelzellen und weitere organspezifische Zellen zur Produktion weiterer
Mediatoren wie Eikosanoiden, Chemokinen, reaktiven Sauerstoffspezies und
Zytokinen. Hauptsächlich zu nennen sind hier TNF-α, IL-1, IFN-γ, IL-6, IL-8,
IL-12 und IL-17. Eine Untergruppe der Zytokine bilden die Chemokine.
Chemokine sind sehr kleine Signalproteine, die man durch die Anordnung von
Cysteinresten im Molekül in verschiedene Klassen einteilen kann. Man
unterscheidet vier Familien, die CC-, die CXC-, die CX3C -und die C-Familie.
Die Funktion der Chemokine kann unterteilt werden in eine
proinflammatorische, induzierbare und eine homöostatische, konstitutive
Wirkung. Im Entzündungsgeschehen dienen sie vor allem der Aktivierung und
Chemotaxis und damit Migration von Leukozyten. Durch ihre Funktion sind
Chemokine bedeutend in der Immunabwehr gegen Infektionen, aber auch in
der Entwicklung von chronischen entzündungsbedingten Krankheiten, wie
Allergien und Autoimmunkrankheiten.
Näher eingegangen werden soll hier nur auf die CC-Chemokine MDC
(macrophage-derived chemokine, CCL22) und TARC (thymus- and activation-
regulated chemokine, CCL17) und ihren Rezeptor CCR4. Sie gehören zu den
Chemokinen, die sowohl proinflammatorisch als auch homöostatisch wirksam
sind, da sie sowohl beteiligt sind an der T-Zell-Entwicklung als auch an der
Page 19
Kapitel 1: Einleitung
11
Rekrutierung von Entzündungszellen durch Chemotaxis. CCR4 wird exprimiert
von dendritischen Zellen, Makrophagen, Basophilen, NK-Zellen und
Thrombozyten, ist aber in erster Linie bekannt durch sein vorwiegendes
Vorkommen auf T-Zellen, vor allem auf dem TH2-Subtyp. Das vermehrte
Vorkommen von CCR4 ist mit vielen Krankheitsbildern verknüpft wie
Lungenfibrose, granulomatöse Lungenerkrankungen und Diabetes. Seine
Rolle in der Sepsis wurde vor allem durch CCR4-/--Mäuse bestätigt. Sie
zeigten ein besseres Überleben nach Induktion einer Sepsis durch operative
polymikrobielle Sepsismodelle oder LPS-Injektion im Vergleich zu septischen
C57BL/6-Mäusen. [30-32].
Die gebildeten Entzündungsmediatoren haben unter anderem Einfluss auf
Blutgefäße und Endothelzellen. Es kommt zur Bildung von Stickstoffmonoxid
und damit zur Vasodilatation und verringertem Blutfluss [33]. Endothelzellen
exprimieren außerdem vermehrt Oberflächenrezeptoren, die eine Adhäsion
der durch die Vasodilatation verlangsamten Leukozyten ermöglichen und
damit die Diapedese zum Entzündungsort vermitteln. Der systemische
Überschuss der Zytokine führt zusätzlich zur Ausprägung klinischer
Symptome. Die Gefäßerweiterung bewirkt einen Blutdruckabfall, außerdem
kommt es zu Fieber, Hyper- oder Hypothermie, Tachykardie, Tachypnoe und
anfangs zu einer Leukozytose. Neben diesen Effekten induzieren Phagozyten
über Präsentation von pathogenen Strukturen die adaptive Immunantwort, die
vermittelt wird von T- und B-Zellen. Zytokine haben hierbei eine zusätzliche
Rolle in der Festlegung der Art der adaptiven Immunantwort. Beispielsweise
differenzieren naive CD4+ T-Helfer-Zellen durch IFN-γ und IL-12 in TH1- und
durch IL-4 in TH2-Zellen. Während TH1-Zellen eine zelluläre Immunantwort über
erhöhte Proliferation zytotoxischer CD8+ T-Zellen auslösen, stimulieren TH2-
Zellen eine humorale Immunantwort über Erhöhung der B-Zell-Proliferation
und Klassenwechsel der gebildeten Antikörper.
Neben der Zytokinproduktion werden durch die eindringenden Mikroben auch
Kaskaden im Plasma aktiviert. Neben dem bereits beschriebenen
Gerinnungssytem wird unter anderem das Komplementsystem aktiviert. Es
führt auf der einen Seite zur direkten Abtötung der Mikroorganismen über die
Bildung eines Membran-Attack-Komplexes, einer Pore in der
Page 20
Kapitel 1: Einleitung
12
Bakterienmembran, oder hilft auf der anderen Seite bei der Beseitigung der
Keime durch Opsonierung.
Diese Überreaktion des Immunsystems kann im Extremfall zu einem
hyperdynamen, septischen Schock bis hin zum multiplen Organversagen
führen. Dann spricht man auch vom MODS (multi organ dysfunction
syndrome).
Über einen langen Zeitraum wurde angenommen, dass die unkontrollierte
Hyperinflammation auf die eindringenden Organismen Ursache für die hohe
Mortalität in der Sepsis sei. Die daraufhin durchgeführten Studien mit dem Ziel
zur Unterdrückung der überschießenden Inflammation schlugen allerdings
fehl. Sowohl TNF-Antagonisten, Il-Antagonisten, als auch Antiendotoxin-
Antikörper zeigten nicht die gewünschten Erfolge. Viele starben trotz
Behandlung, vor allem an Sekundärinfektionen. Der Grund hierfür ist die
zweite Phase der Immunantwort. Nach dem Zytokinsturm kommt es zur
kompensatorischen Gegenregulation. Antiinflammatorische Zytokine wie IL-10
sind diesmal die Hauptakteure. Es kommt zur Apoptose peripherer
Immunzellen, zur Anergie der T-Zellen und zu einem Wechsel der T-Helfer-
Zellen zum TH2-Typ. Dies kann im Extremfall zur Immunparalyse führen und
damit zur starken Anfälligkeit für Sekundärinfektionen [34].
1.3 Sepsismodelle
Da die Sepsis ein sehr komplexes Krankheitsbild darstellt, ist es nicht einfach
die zu Grunde liegenden Mechanismen aufzuklären.
Hierbei dienen Tierexperimente als unverzichtbares Modell zur Untersuchung
der Pathomechanismen im Gesamtorganismus und Bindeglied zwischen
molekular- und zellbiologischen Experimenten und klinischen Studien.
Zur Induktion einer Sepsis wurden im Laufe der Jahre viele Modelle entwickelt
von der Injektion von bakteriellen Bestandteilen bis hin zum operativen
Sepsismodell. Im Folgenden sollen die wichtigsten Vorgehensweisen kurz
erläutert werden.
1.3.1 Exogene Verabreichung von Toxinen und Bakterien
Bakterien und vor allem deren Zellwandbestandteil LPS sind oft Ursache für
die Entstehung einer Sepsis. Als Modell wurde deshalb die Injektion von LPS
Page 21
Kapitel 1: Einleitung
13
entwickelt. Die Applikation erfolgt intravenös, intraperitoneal oder für einen
längeren Zeitraum als Bolus. Neben in vivo-Gabe ist die Stimulation mit LPS
auch in vitro Gang und Gebe um die Effekte auf einzelne Zellpopulationen
beurteilen zu können. Nach Injektion von LPS in die Tiere kommt es zur
Ausbildung der typischen Symptome eines septischen Schocks.
Um zusätzlich die Expansion der Bakterien und deren Elimination durch den
Wirt, was im LPS-Modell nicht betrachtet werden kann, zu berücksichtigen,
können auch lebendige Bakterien injiziert werden. Klarer Vorteil beider
Modelle ist die einfache Durchführung und die kontrollierbare Menge
eindringender Mikroben. Die Ausprägung der Symptome ist kaum durch
experimentelle unbeeinflussbare Variationen zu ändern. Die Übertragbarkeit
beider Modelle auf den Verlauf des septischen Schocks beim Menschen
konnte bis jetzt allerdings noch nicht gezeigt werden. Um dem wirklichen
Krankheitsverlauf in der Sepsis näher zu kommen, wurden operative Modelle
entwickelt, die von einer Peritonitis ausgehen. Die beiden wichtigsten
Methoden sollen folgend kurz erläutert werden.
1.3.2 Cecal Ligation and Puncture (CLP)
CLP ist das wohl am häufigsten verwendete operative Sepsismodell. Dieses
Modell wurde erstmals 1980 beschrieben [35].
Bei dem CLP-Modell wird das Zökum unterhalb der Ileozökalklappe ligiert und
mit einer Nadel punktiert. Die aus dem künstlich erschaffenen Leck
austretenden Darmkeime erzeugen eine Peritonitis. Die Entwicklung der
Symptome von Peritonitis und Sepsis nach Darmperforation benötigt längere
Zeit und ähnelt deswegen stärker einem klinischen Verlauf als nach LPS-
Injektion.
1.3.3 Colon Ascendens Stent Peritonitis (CASP)
Das CLP-Modell wurde in den letzten Jahren weiterentwickelt und führte zu
einem weiteren operativen Modell, der CASP. Erstmals wurde es 1998 von
Zantl et. al. bei der Anwendung an Mäusen beschrieben [36]. Das neue Modell
sollte der generalisierten Peritonitis beim chirurgischen Patienten möglichst
nahe kommen um die immunologische Pathophysiologie einer Sepsis
realitätsnah untersuchen zu können.
Page 22
Kapitel 1: Einleitung
14
Der Maus wird bei der CASP ein Stent in den aufsteigenden Dickdarmteil
implantiert. Dadurch wird eine Verbindung zwischen Darmlumen und
Peritoneum hergestellt und ein stetiger Ausfluss von Darminhalt in den
Bauchraum gewährleistet. Die heraustretenden Bakterien führen zur
Ausbildung einer diffusen Peritonitis und schließlich zur polymikrobiellen
Sepsis.
Die CASP wurde von Niko Zantl im Rahmen einer chirurgisch-
mikrobiologischen Kooperation erarbeitet und 1997 erstmals publiziert. Ziel
der Studien war es, ein Tiermodell in der Maus für die postoperative
abdominelle Sepsis zu entwickeln, um damit die immunologische
Pathophysiologie während des Verlaufs einer Sepsis analysieren zu können.
Durch Variation des Durchmessers von 14G bis 18G bei den verwendeten
Stents kann die Mortalität der CASP beeinflusst werden. Das genaue
operative Verfahren der CASP ist unter 3.2.3 beschrieben.
Page 23
Kapitel 2: Fragestellung der Arbeit
15
2 Fragestellung der Arbeit
Die Bedeutung der Thrombozyten wuchs in den letzten Jahren mehr und mehr
über ihre alleinige Funktion in der Hämostase hinaus. Ziel der vorliegenden
Arbeit ist es, die erweiterte Rolle dieser kernlosen Zellen in der Sepsis zu
untersuchen. Septische Patienten leiden oft unter Thrombozytopenie und
disseminierter intravasaler Gerinnung, die die Prognose der Krankheit
zusätzlich verschlechtern. Die Verhaltensweisen der Thrombozyten in der
Sepsis sollen sowohl in vitro als auch in vivo in einigen Punkten untersucht
werden. Schwerpunkt ist dabei in vitro die Reaktion auf Stimulation mit
Bakterienbestandteilen und in vivo die Verteilung der Thrombozyten im
septischen Mausmodell. Induziert wird das komplexe Krankheitsbild der
Sepsis dabei über die Colon Ascendens Stent Peritonitis (CASP), die eine
Möglichkeit bietet die Situation einer polymikrobiellen Sepsis realistisch
nachzubilden.
Neben den Thrombozyten als ein Akteur in der Sepsis und möglicher
Angriffspunkt für neue therapeutische Strategien, soll auch der
Chemokinrezeptor CCR4 untersucht werden. Da neben Immunzellen auch
Blutplättchen CCR4 exprimieren und CCR4-/--Mäuse ein besseres Überleben
nach CASP zeigen, wird hier ein möglicher Zusammenhang vermutet und am
Mausmodell untersucht [30].
Page 24
Kapitel 3: Material und Methoden
16
3 Material und Methoden
3.1 Material
3.1.1 Reagenzien und Chemikalien
Aceton Merck, Darmstadt
Ammoniumchlorid (NH4Cl) Merck, Darmstadt
Arg-Gly-Asp-Ser (RGDS) Sigma-Aldrich Chemie,
Steinheim
Bovines Serumalbumin ICN Biomedicals, Aurora, USA
Bovine Albumin 22% Ortho-Clinical Diagnostics Inc.
Neckargmünd
Biotin Blocking System, ready-to-use Dako, Glostrup
Cellfix BD Biosciences
Chloroform Sigma-Aldrich Chemie,
Taufkirchen
Complete Roche, Mannheim
DABCO Merck, Darmstadt
Destilliertes Wasser GIBCO, InvitrogenTM
- DNase/RNase frei
Diethylpyrocarbonat (DEPC) Sigma-Aldrich Chemie,
Taufkirchen
DNAse Qiagen, Hilden
dNTPs MBI Fermentas, St. Leon – Rot
EDTA Merck, Darmstadt
Einbettmedium Tissue - Tek® Sakura Finetek Europe B.V.,
Zoeterwoude, Niederlande
Ethanol unvergällt Carl Roth, Karlsruhe
FACS - Puffer Becton Dickinson, Heidelberg
Fötales Kälberserum, inaktiviert Sigma-Aldrich Chemie,
Taufkirchen
FcR Blocking Reagent, mouse Miltenyi Biotec
Hepes Puffer Sigma-Aldrich Chemie,
Steinheim
Page 25
Kapitel 3: Material und Methoden
17
Histopaque -1083 Sigma-Aldrich Chemie,
Steinheim
Isotonische Natriumchlorid-Lösung Delta Select, Pfullingen
Korsolex Bode Chemie GmbH & Co. KG,
Hamburg
Lipopolysaccharid (LPS), E.Coli O127:B8 Sigma-Aldrich Chemie,
Taufkirchen
Lyse-Puffer BD Biosciences
Mercaptoethanol Sigma-Aldrich Chemie,
Taufkirchen
N - Acetyl - L - alanyl - L - Glutamin Biochrom AG, Berlin
Natriumhydroxid (NaOH) Merck, Darmstadt
PBS Dulbecco w/o Ca2+, Mg2+ Biochrom AG, Berlin
PBS, steril Sigma-Aldrich Chemie,
Taufkirchen
PBS Dulbecco (1x) w Ca2+, w Mg2+ Biochrom AG, Berlin
Pefa - Bloc Roche, Mannheim
Peroxidase Blocking Reagent (Dako) Dako
Phenylmethylsulfonylfluorid (PMSF) Sigma-Aldrich Chemie,
Taufkirchen
2-Propanol Sigma-Aldrich Chemie,
Taufkirchen
Random Primers Invitrogen, Karlsruhe,
SuperArrays
RNase ZAP® Sigma-Aldrich Chemie,
Taufkirchen
RNAsin Ribonuclease Inhibitor Promega, Mannheim
Superscript II RNAse H
Reverse Transcriptase Invitrogen, Karlsruhe
TaqMan® Universal PCR Mastermix Applied Biosystems,
Branchburg
TaqMan Assay Reagents
- für Real – Time PCR Applied Biosystems,
Branchburg
Page 26
Kapitel 3: Material und Methoden
18
Tergitol Sigma-Aldrich Chemie,
Taufkirchen
TRAP-6, PAR-1 (1-6) (human) BACHEM, Bubendorf, Schweiz
Triton X 114 Sigma- Aldrich Chemie,
Steinheim
TRI Reagent Sigma-Aldrich Chemie,
Taufkirchen
Türck`s Lösung Merck, Darmstadt
Tween 20 Sigma-Aldrich Chemie,
Taufkirchen
3.1.2 Kits
CD4+ T Cell Isolation Kit Miltenyi Biotec GmbH
RNeasy® Mini Kit Qiagen, Hilden
TSATM Biotin System, für 200-600 Proben PerkinElmer, Wartham , USA
3.1.3 Primer
murine GAPDH probe (Farbstoff: VIC) Applied Biosystems,
Branchburg
CCR4 TaqMan assay (Mm99999052_s1) Applied Biosystems,
Branchburg
3.1.4 Antikörper
Rat polyclonal IgG, PE-labeled (P190-2) emfret analytics Eibelstadt
Rat polyclonal IgG, FITC-labeled (P190-1) emfret analytics Eibelstadt
rat monoclonal anti-mouse GPIbβ (CD42c)
FITC labeled (M050-1)
emfret analytics Eibelstadt
donkey polyclonal anti-goat IgG
PE-labeled (705-708-125)
Rockland, Gilbertsville
Purified rat anti-mouse CD62P (550289) BD PharmingenTM
Purified Rat anti-mouse Ly-6G and Ly-6c
(553123)
BD PharmingenTM
Page 27
Kapitel 3: Material und Methoden
19
FITC labeled monoclonal anti-mouse CD4
(130-091-608)
Miltenyi Biotec
PE conjugated anti-mouse CD45RB
(120455)
eBioscience
rat monoclonal anti-mouse CD34
PE-labeled (12-0341)
eBioscience
PE-CyTM5 Mouse Anti-Human CD62P
(551142)
BD PharmingenTM
FITC PAC-1 (340507) BD PharmingenTM
CD42a-PE (558819) BD PharmingenTM
IgG1-PE-Cy5 (555750) BD PharmingenTM
PE-Cy5 monoclonal anti-human CD41,
Klon P2 (6607116)
Beckman Coulter GmbH
Isotyp Tricolor V, PNIM 1672 Beckman Coulter GmbH
POD conjugated F(ab’)2 Fragment mouse
anti-ratIgG (H+L), (212-036-168)
Jackson
Immuno Research
Farbstoffe
Alexa 488 (A-11090) Molecular Probes
Streptavidin TRITC (016-030-084) Dianova
3.1.5 Puffer und Lösungen
Erythrozyten - Lysepuffer: Aqua bidest
155 mM NH4Cl
10 mM KHCO3
0,1 mM EDTA
Sucrose-Lösung 15 ml HEPES-Waschpuffer
6 % Sucrose
0,06 % Triton X114
Lösung A 80 g NaCl
Page 28
Kapitel 3: Material und Methoden
20
2 g KCl
10 g NaHCO3
0,5 g NaH2PO4×H2O
500 ml Aqua bidest
Tyrode Ansatz 2 ml Glucose 10% in A. bidest
5 ml Lösung A
1 ml HEPES 0,5M pH 7,3
40 mg Adenosin
18 mg Theophyllin
92 ml A. bidest
3.1.6 Laborgeräte
ABI Prism® 7000 Sequence Detection
System
Applied Biosystems, Foster
City
Durchflusszytometer Beckman Coulter,
FC 0
Beckman Coulter, Krefeld
Digitalkamera Power Shot S50 Canon Deutschland GmbH,
Krefeld
Digitalwaage BL 150 S Sartorius AG, Göttingen
Eismaschine IceLine KF85
Fluoreszenzmikroskop Leica DM IL
Migel
Leica Microsystems, Wetzlar
Halbmikroküvetten Brand, Wertheim
Magnetrührer (MR 3001 K)
Magnetständer MACS® MultiStand
Heidolph, Schwabach
Miltenyi Biotec,
Bergisch Gladbach
Mikrotom - Kryostat Cryo - Star,
HM 560
Microm, Walldorf
NanoDrop 2000/2000c
pH-Meter inoLab
Peqlab Biotechnologie GmbH,
WtW, Weilheim
Pipetten Eppendorf, Hamburg
Reagenzglasschüttler Vortex Merck Eurolab N.V., Leuven,
Belgien
Spectrophotometer NanoDrop 2000,
peqLab
Biotechnologie GmbH,
Erlangen
Page 29
Kapitel 3: Material und Methoden
21
Sterilwerkbank Microflow Nunc GmbH & Co. KG,
Wiesbaden
Sterilwerkbank Herasafe Heraeus, Hanau
Thermocycler Biometra, Göttingen
Ultraschallbad Sonorex TK 52 Bandelin, Berlin
Zellzählung Sysmex KX-21N Sysmex Digitana AG Horgen
Zentrifugen:
Centrifuge 5415 R Eppendorf, Hamburg
Megafuge 1.0 R Heraeus, Hanau
Cytospin 4 Shandon
Chirurgisches Besteck:
Pinzette Asanus, Neuhausen ob Eck
Nadelhalter, Mikronadelhalter Asanus, Neuhausen ob Eck
Schere, Mikroschere Asanus, Neuhausen ob Eck
3.1.7 Verbrauchsmaterialien
Chirurgisches Nahtmaterial:
Mariderm schwarz 7/0 USP Catgut GmbH, Markneukirchen
Polyester weiß 4/0 USP Catgut GmbH, Markneukirchen
96-Well Optical Raction Plate
Deckgläser
Applied Biosytems, Foster City, USA
Superior, Marienfeld
Einbettschälchen Cryomold® Sakura Finetek Europe B.V., Zoeterwoude,
Niederlande
Einmalskalpelle Ansell Medical, Cergy Pontoise, Frankreich
Einmalspritzen 1ml Dispomed, Gelnhausen
Einmalspritzen Omnican® F 1 ml Braun, Melsungen
Einmalspritze 5 ml Injekt Braun, Melsungen
Halbmikroküvetten Brand, Wertheim
Hämatokritkapillaren Brand, Wertheim
Kryoröhrchen Nunc, Roskilde, Dänemark
MicroAmpTM,
Optical Adhesive Film
Applied Biosystems, Branchburg, USA
Page 30
Kapitel 3: Material und Methoden
22
Objektträger Superfrost® Plus Menzel Gläser, Braunschweig
Optische Platten
(Real – Time PCR)
Applied Biosystems, Branchburg, USA
PCR Softtubes 0,2ml
Pipettenspitzen
Biozym Scientific GmbH, Oldendorf
Eppendorf, Hamburg
Reaktionsgefäße Eppendorf, Hamburg
Röhrchen für FACS - Analyse Becton Dickinson, Heidelberg
Safe Seal-Tips® professional
Spritzenvorsatzfilter
0,2µm, 0,45µm
Biozym
Merck, Bruchsal
Venenverweilkanülen 16G BD Vialon Biomaterial, Heidelberg
Wattestäbchen care&serve
Zellsiebe 70 µm BD Pharmingen, Heidelberg
Zentrifugenröhrchen:
Spitzboden 15 ml und 50 ml Greiner Bio-One, Frickenhausen
Rundboden 12 ml Greiner Bio-One, Frickenhausen
3.2 Methodik in vivo
3.2.1 Genehmigung der Tierversuche
Gemäß dem deutschen Tierschutzgesetz wurden die Tierversuche vor der
Durchführung von der zuständigen Behörde, dem Landesveterinäramt und
dem Lebensmitteluntersuchungsamt Mecklenburg-Vorpommern genehmigt,
gemäß Aktenzeichen LALLF M-V/TSD/7221.3-11-025/08.
3.2.2 Tiere und Tierhaltung
Als Versuchstiere wurden sowohl C57BL/6 Mäuse als auch CCR4-/- Mäuse
verwendet. Beide Gruppen waren weiblich und bei Versuchsdurchführung
8-12 Wochen alt mit einem Gewicht von 20-25g. Die C57BL/6 Mäuse wurden
von Charles River Laboratories, Research Models and Services, Germany
GmbH in Sulzfeld Frankreich bezogen und die knock-out Mäuse stammen aus
eigener Zucht. Gehalten wurden die Tiere in Makrolonkäfigen (Ehret GmbH
Emmendingen,Deutschland) mit Gruppengrößen von fünf bis zehn Tieren. Der
Käfig war ausgestattet mit 200g Einstreu (Lignocel®, Naturholzfasern), Wasser
Page 31
Kapitel 3: Material und Methoden
23
ad libitum und Alleinfutter in pellettierter Form (M-Z Extrudient, ssniff
Spezialdiäten GmbH). Die Tiere wurden im Biotechnikum in der Walther-
Rathenau-Straße, Greifswald, gehalten, das durch die Ernst-Moritz-Arndt-
Universität Greifswald angemietet wird. Zur Gewährleistung gleicher
Versuchsbedingungen wurden die Tiere eine Woche vor Durchführung der
Experimente unter gleichen Bedingungen im Tierstall eingewöhnt. Alle
Tierversuche wurden unter Einhaltung des deutschen Tierschutzgesetzes
durchgeführt.
3.2.3 Tiermodelle
Zur Sepsisinduktion wurde das Modell der Colon Ascendens Stent Peritonitis
(CASP) angewendet.
3.2.3.1 Anästhesie
Zur Narkotisierung der Versuchstiere wurde eine Injektionlösung aus
2% Rompun, 25 mg/ml Ketanest und 0,9% NaCl frisch hergestellt und 10 µl
pro Gramm Körpergewicht intraperitoneal injiziert. Ungefähr 5 Min nach der
Injektion wurde die Tiefe der Narkose durch Setzen eines Schmerzreizes an
den Hinter- und Vorderpfoten überprüft und gegebenenfalls 0,1 ml
nachgespritzt. Im narkotisierten Zustand wurde die Maus rücklings für die
Operation auf einer Styroporplatte an Vorder- und Hinterläufen fixiert.
3.2.3.2 Colon Ascendens Stent Peritonitis (CASP)
Bei der CASP wird durch einen kontinuierlichen Austritt von Bakterien der
natürlichen Darmflora in die Bauchhöhle eine disseminierte Peritonitis und
Sepsis hervorgerufen. Der Übertritt der Pathogene wird durch Einbau eines
Stents in das Colon Ascendens der Maus ermöglicht. Der Stent verbindet
Darmlumen und Bauchhöhle und sorgt für einen Übertritt von Fäzes in den
Bauchraum. Je nach Größe des eingesetzten Stents (14G, 16G, 18G) variiert
die Schwere der Sepsis und damit die Letalität. In dieser Arbeit wurden
ausschließlich Stents mit einem Durchmesser von 16 Gauge zur
Sepsisinduktion verwendet.
Page 32
Kapitel 3: Material und Methoden
24
Folgende Instrumente wurden verwendet:
- Mikrochirurgisches Besteck: Mikronadelhalter, Pinzette, Mikroschere
- Nadelhalter
- Schere
- Wattestäbchen, Tupfer, Pflaster
- Venenverweilkatheter: 16G VenflonTM, 1,7x45 mm; 180 ml/Min
(Becton Dickinson, Heidelberg)
- Faden: 4/0 Polyester (Catgut GmbH, Marktneukirchen)
7/0 Mariderm schwarz (Catgut GmbH, Marktneukirchen)
Durchführung der Operation
Vor der Operation wird das Versuchstier wie in 3.2.3.1 beschrieben
anästhesiert. Zur Vorbereitung wird der 16G Venenverweilkatheter etwa 2 mm
nach Beginn der Kunststoffummantelung mit einer Schere zirkulär eingeritzt.
Die entstandene Inzision im späteren Dickdarmstent dient dann als Widerlager
für den Faden.
Nachdem die Maus ausreichend narkotisiert wurde, wird sie in Rückenlage auf
einer Styroporplatte mit Pflasterstreifen an den Rück- und Vorderläufen fixiert.
Das Bauchfell wird desinfiziert mit 70%igem Ethanol und mit einem Tupfer
glatt gestrichen um zu vermeiden, dass Haare in die Bauchhöhle gelangen.
Die desinfizierte Bauchhaut wird mit einer Pinzette leicht angehoben um die
folgende Inzision der Haut zu erleichtern. Die Haut und analog die darunter
liegende Muskelfaszie werden ab der Symphyse unter Vermeidung von
Blutung ca. 15 mm nach kranial inzidiert.
Mittels zweier Wattestäbchen wird der Zökalpol aufgesucht um anschließend
Zökum, Colon ascendens und terminales Ileum vorsichtig nach außen zu
luxieren. Das Colon ascendens wird ca. 15 mm nach der Mündung des
terminalen Ileums mit Mariderm 7/0 durchstochen und ein Knoten wird
vorgelegt. Nun erfolgt die Punktion der antimesenterialen Seite des Colon
ascendens mit dem vorbereiteten Venenverweilkatheter unmittelbar aboral
des vorgelegten Knotens. Der Katheter wird anschließend vorsichtig vorwärts
geschoben, bis sich die Kerbe des Plastikmantels auf Höhe der Serosa
befindet. Der Stent wird nun mit den freien Enden des Fixierungsfadens
mittels chirurgischen Knoten in der Inzision befestigt und so am Hinausgleiten
Page 33
Kapitel 3: Material und Methoden
25
gehindert. Eine weitere Fixationsnaht erfolgt von hier aus nach aboral durch
einen Stich durch die Darmwand. Zuletzt wird die Nadel vorsichtig
herausgezogen und der Stent auf ca. 1 mm über der Serosa gekürzt.
Vor der Rückverlagerung der Darmteile in die Bauchhöhle, wird der Stent
durch leichten Druck mit zwei Wattestäbchen auf das Zökum mit Fäzes gefüllt.
Sind Zökum, Colon ascendens und terminales Ileum reponiert, werden ca.
0,5 ml 0,9%iger Kochsalzlösung in die Bauchhöhle injiziert um den
Flüssigkeitsverlust während der OP auszugleichen. Die Wunde wird
fortlaufend mit 4/0 Polyesterfaden vernäht, sodass sowohl Peritoneum als
auch Muskulatur und Haut gut verschlossen sind.
3.2.3.3 Schmerztherapie
Alle Mäuse erhielten postoperativ eine Schmerztherapie mit Buprenorphin
0,1 mg/kg pro Tag i.m..
3.2.4 Organentnahme
Vor Entnahme der einzelnen Organe wurden die Tiere wie unter 3.2.3.1
beschrieben narkotisiert. Begonnen wurde mit der Punktion des
Retroorbitalplexus des Auges um Blut zu gewinnen. Eine mit EDTA
beschichtete Mikro-Hämatokrit-Kapillare wurde genutzt zur Unterbindung der
Blutgerinnung. Etwa 1 ml Blut wurde aufgefangen und durch bereits im Gefäß
vorgelegtes Antikoagulanz (170 µl ACDA und 5 µl Hirudin) antikoaguliert. Das
Blut wurde bis zur Weiterverarbeitung ruhig und bei RT gelagert. Zur weiteren
Organentnahme wurde das Tier in Rücklage fixiert und nach Desinfektion der
Haut an der Operationsnaht bzw. Linea alba mithilfe einer Mikroschere
vollständig eröffnet. Milzen zur CD4+-Zellisolation wurden vorsichtig
freipräpariert, entnommen und in PBS auf Eis gelagert. Organe für die
histologische Aufarbeitung wurden in Tissue-Tek® gebettet und in flüssigem
Stickstoff schockgefroren. Entnommen wurden linker Leberlappen, Milz, linke
Niere und die Lunge. Bei der Entnahme der Lunge wurde vorher die Trachea
durchtrennt um über diese Öffnung beide Lungenlappen mit Tissue-Tek® zu
füllen. Dadurch wurde der Erhalt der feinen Alveolenstruktur trotz des
Schockgefrierens gewährleistet. Bis zum Schneiden der Organe wurden sie
bei -80°C gelagert.
Page 34
Kapitel 3: Material und Methoden
26
3.3 Zellbiologische Methoden mit humanen Zellen
3.3.1 Abnahme und Präparation der Blutproben
Die Blutproben zur Durchführung der Stimulationsexperimente wurden von
gesunden, freiwilligen Spendern des Instituts für Immunologie und
Transfusionsmedizin Greifswald, denen im Rahmen einer Blutspende Vollblut
abgenommen wurde, gewonnen. Das kubital, venös entnommene Blut wurde
mit Citrat antikoaguliert und ruhig gelagert um eine Aktivierung von
Thrombozyten zu vermeiden. Anschließend folgte die Stimulation der Zellen.
Folgende Stimulanzien wurden sowohl für Vollblut als auch für
plättchenreiches Plasma verwendet:
- Lipopolysaccharid (LPS) aus E.Coli O127:B8, Sigma-Aldrich Chemie,
Taufkirchen
- TRAP-6, PAR-1 (1-6) (human) (thrombin receptor agonist peptide-6),
Bachem
3.3.2 Stimulation von Vollblut
Humanes Citratvollblut wurde vor dem Versuch mit dem Sysmex Cellcounter
gezählt. Pro FACS-Röhrchen wurden 100 µl Vollblut vorgelegt und entweder
mit 1-10 µg/ml LPS (E.coli O127:B8), 50 µmol/l TRAP-6 oder beidem
stimuliert. Die Negativkontrolle wurde mit 1 µl PBS (+Ca/+Mg) versetzt.
Anschließend wurden die Proben 4 h bei 37°C inkubiert. Daran schloss sich
die Messung der Oberflächenexpression der Aktivierungsmarker mittels
Durchflusszytometrie.
3.3.3 Stimulation von plättchenreichem Plasma (PRP)
Das Citratvollblut wurde 20 Min. bei 860 rpm, ohne Bremse zentrifugiert zur
Gewinnung des thrombozytenreichen Plasmas. Das Plasma wurde vorsichtig
in ein neues FACS-Röhrchen überführt und die Anzahl der Thrombozyten
wurde mit dem Sysmex Cellcounter bestimmt. Die Probe wurde mit PBS
(+Ca2+, +Mg2+) auf eine Zellkonzentration von 1x108/ml eingestellt. Im
Page 35
Kapitel 3: Material und Methoden
27
Testansatz wurden pro Röhrchen dann 50 µl PRP überführt, was einer Anzahl
von 5x106 Plättchen entsprach. Zur Stimulation wurden jeweils LPS
(10 µg/ml), TRAP-6 (50 µmol/l), beides oder als Negativkontolle PBS
hinzugesetzt und die Proben für 2-4 h bei 37°C inkubiert. Anschließend
erfolgte die Messung der Aktivierung mittels Markierung durch spezifische
Antikörper in der Durchflusszytometrie.
3.4 Zellbiologische Methoden mit murinen Zellen
3.4.1 Isolation muriner Thrombozyten
Bevor mit der eigentlichen Isolation begonnen wurde, wurde zum Tyrode
Ansatz sowohl Hirudin (1000 U/ml, 4 µl auf 10 ml Tyrode Ansatz) als auch
BSA (10%ig) hinzugefügt. Die Lösung wurde auf 37°C erwärmt und mit
25%iger HCl auf einen pH-Wert von 6,5 eingestellt.
Dem antikoagulierten murinen Blut wurde 1 ml des angesetzten Tyrode-
Ansatzes hinzugefügt. Schütteln oder vortexen des Eppendorfgefäßes wurden
unterlassen um eine Aktivierung der Thrombozyten zu vermeiden. Die
Mischung wurde bei 200xg 5 Min. ohne Bremse bei RT zentrifugiert. Das nun
als obere Schicht vorliegende PRP wurde vorsichtig in ein neues
Eppendorfgefäß überführt und nochmal mit 1 µl ProstaglandinE1/ml
antikoaguliert. Bei 169xg ohne Bremse wurden für 5 Min. die Erythrozyten
abzentrifugiert. Das PRP wurde wieder in ein neues Eppendorfgefäß überführt
und mit 111 µl ACDA/ml versetzt. 100 µl des PRP wurden zur Zellzählung
verwendet, der Rest wurde zur weiteren Verwendung bei 1400xg 5 Min. bei
RT pelletiert.
3.4.2 Aufarbeitung muriner Lymphozyten
3.4.2.1 Isolation von PBMCs aus dem peripheren Blut
Die Isolation der Lymphozyten und Monozyten aus dem peripheren murinen
Blut erfolgte mittels Zentrifugation über einen Dichtegradienten. Hilfsmittel war
hierbei HISTOPAQUE 1083, eine Lösung aus Polysaccharose und
Natriumdiatrizoat mit einer Dichte von 1.083 g/ml.
Das Blut wurde mit PBS 1:2 verdünnt. Histopaque (3 ml) wurde vorgelegt und
dann vorsichtig mit 3 ml des vorverdünnten Blutes überschichtet.
Anschließend wurde 30 Min. bei RT und 400xg ohne Bremse zentrifugiert.
Page 36
Kapitel 3: Material und Methoden
28
Durch die Zentrifugation werden die Blutzellen separiert. Die Erythrozyten
aggregieren mit der Polysaccharose des Histopaques und befinden sich dann
zusammen mit den Granulozyten aufgrund ihrer hohen Dichte in der untersten
Schicht. Die mononukleären Zellen liegen als Zellring auf der Histopaque-
Phase. Die darüber befindliche Plasmaschicht wurde bis ca. 5 mm über den
PBMCs abgenommen und verworfen. Die PBMCs wurden überführt und mit
PBS verdünnt. Es folgten Waschschritte zur Elimination überschüssiger
Thrombozyten und zur vollständigen Entfernung des Histopaques. Zuerst
wurden die Zellen zentrifugiert bei 250xg für 10 Min. Der entstehende
Überstand wurde abgegossen und das Röhrchen wurde wieder mit 5 ml PBS
aufgefüllt, vermischt und bei 250xg 10 Min. zentrifugiert. Dieser Waschritt
wurde wiederholt und anschließend wurde das Zellpellet in 500 µl PBS
aufgenommen und mit dem Sysmex Zellcounter gezählt. Zur RNA-Isolierung
wurden die Zellen 10 Min. bei 250xg pelletiert und bei -80°C gelagert.
3.4.2.2 Isolation von CD4+-Zellen aus der Milz
Die Isolation der T-Helfer-Zellen aus der Milz erfolgte über ein indirektes
magnetisches Markierungssystem, bei dem alle Nicht-CD4+-Zellen depletiert
werden. Alle unerwünschten Milzzellen werden mithilfe eines Cocktails aus
Biotin-konjugierten Antikörper markiert und bleiben später auf einer
magnetischen Säule hängen, so dass im Eluat die gewollten CD4+-Zellen sind.
Zuvor muss die Milz zu einer Einzelzellsuspension aufgearbeitet werden.
Während des gesamten Isolationsprozesses wurden die Milzen gekühlt und
die Waschschritte erfolgten bei 4°C für 10 Min. bei 300xg in der Zentrifuge.
Während des gesamten Vorgangs wurden alle Puffer gekühlt verwendet. Die
Inkubationen wurden bei 4°C durchgeführt um unspezifische Zellmarkierungen
zu vermeiden. Die Milz wurde durch ein 70 µm Zellsieb gedrückt und dabei mit
PBS/2%FCS (fötales Kälberserum) gespült. Danach wurden die Zellen
gewaschen und das Zellpellet wurde in PBS resuspendiert. Anschließend
wurden die Erythrozyten lysiert. Die Lyse wurde durch Zugabe von
PBS/2%FCS abgestoppt. Nach einem weiteren Waschschritt wurde das Pellet
in 1 ml PBS/2%FCS aufgenommen und am Sysmex Cellcounter wurden die
Zellen gezählt. Die Zellen wurden nun nochmal bei 470xg 5 Min. zentrifugiert,
um mit dem magnetischen Labeling zu beginnen. Das Pellet wurde in MACS-
Page 37
Kapitel 3: Material und Methoden
29
Puffer resuspendiert (40 µl pro 1x107 Zellen) und mit dem Biotin-Antikörper-
Cocktail für 10 Min. bei 4°C versetzt (10 µl pro 1x107 Zellen),. Der Cocktail
enthält Antikörper gegen CD8a, CD11b, CD11c, CD19, CD45R (B220),
CD49b (DX5), CD105, Anti-MHC-class II, und Ter-119 und markiert damit
CD8+-T-Zellen, B-Zellen, dendritische Zellen, Makrophagen, NK-Zellen und
Granulozyten. Nach weiterer Zugabe von MACS-Puffer (40 µl pro 1x107
Zellen) wurden die Anti-Biotin MicroBeads dazu pipettiert und 15 Min.
inkubiert. Alle Zellen, die durch einen Antikörper markiert werden, binden
zusätzlich die magnetischen Microbeads und verbleiben dadurch später an
der Säule. Die Zellen wurden mit 1 ml MACS-Puffer gewaschen und
zentrifugiert für 10 Min. bei 300xg und dann in 500 µl Puffer aufgenommen.
Für die Separation wurden die MACS Separation Colums in dem Magneten in
Position gebracht und mit einem Pre-Separation-Filter versehen um eine
Verklumpung durch zu große Zellreste in der Säule zu vermeiden. Bevor die
Probe aufgetragen wurde, wurden beide Systeme mit 500 µl Puffer gespült.
Nach dem Durchlauf der Probe wurde dreimal mit 500 µl Puffer nachgespült.
Vor jedem Schritt wurde darauf geachtet, dass die Flüssigkeit vorher komplett
die Säule passiert hat. Die isolierten Zellen wurden gezählt und zur
Überprüfung der Reinheit mit einem CD4+-FITC und einem CD45-PE
markierten Antikörper durchflusszytometrisch gemessen. Für die RNA-
Isolation wurden die Zellen bei 300xg 10 Min. zentrifugiert.
3.5 Molekularbiologische Methoden
3.5.1 RNA-Isolation
Die RNA-Isolation erfolgte sowohl mit TRI Reagent als auch mit dem RNeasy
Mini Kit. Vor der Isolation wurde der Arbeitsplatz mit RNase ZAP® von RNasen
befreit. Zur Isolation wurden ausschließlich Pipetten mit Filter Tips verwendet.
RNA-Isolation mit TRI Reagent
Die Gesamt-RNA aus den PBMCs wurde mit der TRIZOL-Methode gewonnen,
modifiziert nach Chomczynski und Sacchi. Nachdem 5-6x106 Zellen isoliert
und pelletiert wurden, wurde 1 ml Trizol dazu gegeben und resuspendiert bis
sich die Zellen gelöst hatten. Durch diese Mischung aus Phenol und
Guanidinisothiozyanat werden Zellbestandteile gelöst und zerstört, während
Page 38
Kapitel 3: Material und Methoden
30
die Integrität der RNA gewährt wird. Nach 5 Min. Inkubation bei RT wurden
200 µl Chloroform dazugegeben und 15 s geschüttelt. Die Probe wurde 7 Min.
inkubiert und 15 Min. bei 12000xg zentrifugiert, wodurch es zur Aufteilung der
Probe in drei Phasen kommt. In der oberen wässrigen Phase befindet sich die
RNA während die untere Protein enthält. Die DNA verteilt sich auf die untere
und mittlere Phase.
Nach Abnahme der oberen Phase, wurde mit 500 µl Isopropanol die RNA
präzipitiert und nach 10 Min. Inkubation bei 12000xg 10 Min. zentrifugiert. Je
nach RNA-Gehalt bildet sich ein gelartiges Pellet. Die RNA wurde noch einmal
mit 75%igen Ethanol gewaschen und anschließend bei RT getrocknet. Zuletzt
wurde die RNA in 20 µl RNase freiem Wasser gelöst und bei -80°C gelagert.
Zur Überprüfung der RNA-Qualität wurden 2 µl der RNA auf den NanoDrop
2000/2000c gegeben.
RNA-Isolation mit dem RNeasy Mini Kit (Quiagen)
Die RNA-Isolation muriner CD4+ Milzzellen erfolgte mit dem RNeasy Mini Kit.
Die Milzzellen wurden nach der Isolation pelletiert und aus maximal 1x107
Zellen wurde entsprechend der Herstellerangaben die RNA isoliert. Zwischen
den einzelnen Schritten wurde ein DNA-Verdau eingefügt. Es wurden 10 µl
DNase und 70 µl des zugehörigen RDD-Puffers auf die mit der Probe
benetzten Säule aufgetragen und 15 Min. bei RT inkubiert. Anschließend
wurde mit dem RW1-Puffer wie angegeben fortgefahren. Zum Schluss wurde
die RNA mit 40 µl RNase-freiem Wasser nach 5 Min. Inkubation von der Säule
bei 15700xg gelöst und bis zur Weiterverarbeitung bei -80°C gelagert.
RNA-Kontrolle mit dem NanoDrop 2000/2000c
Nach RNA-Isolation wurden 2 µl der Probe auf den NanoDrop gegeben und
photometrisch untersucht. Über die Absorption bei 260 nm wurde die
Konzentration der RNA bestimmt. Mit dem Quotienten 260 nm/280 nm wurde
die Proteinverunreinigung ermittelt, da Protein Licht einer Wellenlänge von
280 nm absorbiert. Es wurde nur RNA weiterverwendet, deren Quotient
zwischen 1,7-2,1 lag und somit eine geringe Verunreinigung aufwies.
Page 39
Kapitel 3: Material und Methoden
31
3.5.2 Reverse Transkription und Polymerasekettenreaktion
Zur Quantifizierung der CCR4-mRNA und damit der Transkriptionsrate der
Zellen wurde eine quantitative Echtzeit-PCR durchgeführt. Hierbei werden die
gewonnenen Nukleinsäuren nach der Polymerasekettenreaktion vervielfältigt
und gleichzeitig eine Quantifizierung mittels Fluoreszenzfarbstoffen
ermöglicht. In dieser Arbeit wurde mit TaqMan Sonden gearbeitet, die am 3‘-
Ende mit einem Quencher-Farbstoff und an 5‘-Ende mit dem Reporter-
Fluoreszenzfarbstoff FAM markiert werden.
Reverse Transkription
Vor der PCR muss die isolierte RNA in cDNA umgewandelt werden. Hierzu
wurde ein Mastermix aus allen benötigten Reagenzien vorbereitet.
Für 1 µg RNA wurde benötigt:
- 2 µl Random Primer
- 8 µl 1st strand buffer
- 4 µl 0,1 M DTT
- 4 µl 10 mM dNTP
- 1 µl RNAsin
- 2 µl Superscript
- 3 µl RNase-freies Wasser
Es wurde mit Handschuhen und auf Eis gearbeitet. Außerdem wurden Filter
Tips als Pipettenspitzen verwendet um Kontamination mit RNase und DNase
möglichst auszuschließen. Nachdem 40 µl Probe bestehend aus Mastermix
und RNA in die PCR Softtubes gegeben wurde, wurde mit dem Thermocycler
cDNA synthetisiert.
Folgendes Programm wurde verwendet:
- 10 Min. 25°C
- 60 Min. 37°C
- 5 Min. 95°C
- 5 Min. 4°C
Eine Probe bestehend aus Mastermix und Wasser statt RNA dient später als
Negativkontrolle.
Page 40
Kapitel 3: Material und Methoden
32
Quantitative Echtzeit PCR
Für die PCR wurde ein PCR-Mastermix hergestellt bestehend aus Mastermix
und Nukleotidsonden.
Der PCR-Mastermix wurde wie folgt hergestellt:
- 12,5 µl TaqMan Mastermix
- 1,25 µl CCR4-FAM (Nukleotidsonde) oder GAPDH-VIC (endogene
Kontrolle)
- 6,25 µl RNase-freies Wasser
20 µl des PCR-Mastermixes wurden pro Well der 96-Well-Platte vorgelegt. Pro
Well wurden jeweils 5 µl cDNA dazugeben. Jede Probe wurde als
Dreifachbestimmung aufgetragen. Die Negativkontrolle wurde einfach
aufgetragen. Die Platte wurde mit einer Klebefolie verschlossen und bei
2800 rpm 3 Min. zentrifugiert. Anschließend wurde sie in den Thermocycler
(Sequence Detection System) überführt und dem Standardprogramm
unterzogen.
Folgendes Programm wurde verwendet:
- 2 Min. 50°C
- 10 Min. 95°C
- 15 s 95°C
- 1 Min. 60°C
Die Auswertung erfolgte mit ABIPrism nach der ΔΔCt-Methode. Der Ct-Wert
gibt den Zyklus der PCR an, an dem das Fluoreszenzsignal des markierten
Primers das erste Mal die Hintergrundfluoreszenz überschreitet. Je niedriger
der Ct-Wert, desto stärker wird das Zielgen also exprimiert.
3.6 Proteinbiochemische Methoden
3.6.1 Durchflusszytometrie humaner Zellen
Zur Untersuchung der Expression von Oberflächenmolekülen auf bestimmten
Zellen in humanem Vollblut oder plättchenreichem Plasma wurde die Methode
der Durchflusszytometrie verwendet. Hierbei werden die Zellen mittels einer
Page 41
Kapitel 3: Material und Methoden
33
Kapillare angesaugt und passieren im Gerät einzeln einen Laserstrahl.
Anhand ihrer Emission von optischen Signalen nachdem sie den Laser
passiert haben, können die Zellen sortiert werden. Neben Größe und
Granularität kann die Expression von Oberflächenmolekülen mithilfe von
gebundenen Fluoreszensfarbstoffen nachgewiesen werden. Im Folgenden
wird die Durchführung der Vollblut- und Plasmaversuche dargestellt. Humanes
Vollblut bzw. PRP wurde mit TRAP-6, einem synthetischen Thrombin-
Analoga, und LPS versehen und die Änderung von Aktivierungsmarkern
PAC-1 und P-Selektin sowie die Aggregatbildung von Thrombozyten mit
Leukozyten wurde untersucht.
3.6.1.1 Vollblutfärbung für die Durchflusszytometrie
Die Blutproben wurden wie unter 3.3.2 beschrieben behandelt. Nach 4 h
Inkubation der Stimulanzien bei 37°C wurden pro 100 µl Probe je 5 µl
Antikörper hinein gegeben. Die Proben wurden anschließend für weitere 15
Min. inkubiert.
Folgende Antikörper wurden unverdünnt verwendet:
- Isotype Tricolor V, Beckman Coulter GmbH
- PE-Cy5 monoclonal anti-Human CD41, Beckman Coulter GmbH
- PE conjugated anti-mouse CD45RB, eBioscience
- PE-CyTM5 Mouse Anti-Human CD62P, BD PharmingenTM
Anschließend wurde pro Röhrchen 3 ml Lysereagenz (BD, 1:10 verdünnt in
Aqua bidest) dazugegeben und mit Hilfe eines Vortexers vermischt. Die Lyse
wurde 7 Min. im Dunkeln inkubiert. Nach 7 Min. Zentrifugation bei 1300 rpm
wurde mit 3 ml PBS/BSA 0,2% gewaschen. Die Messung erfolgte innerhalb
von 4 h am FC500 der Firma Beckmann-Coulter unter Verwendung eines
geeigneten FACS-Messprotokolls.
Page 42
Kapitel 3: Material und Methoden
34
3.6.1.2 Färbung des PRP
Das PRP wurde wie unter 3.3 beschrieben gewonnen und stimuliert. Als
alternative „Isotyp-Kontrolle“ für den PAC-1 Antikörper wurden zuerst 5 µl
RGDS (10 mg/ml) dazupipettiert und die Proben jeweils für 10 Min. bei RT
inkubiert. Das Tetrapeptid Arg-Gly-Asp-Ser (RGDS) bindet einen Teil des
aktivierten Fibrinogenrezeptor GPIIb/IIIa auf Thrombozyten und blockiert dort
spezifisch die Bindungsstelle für PAC-1 [37]. Es wird in den Herstellerangaben
des Antikörpers als Isotypkontrolle angegeben. Danach wurden 5 µl der
angegebenen Antikörper hinzu pipettiert und die Proben nochmals für 10 Min.
bei RT inkubiert.
Folgende Antikörper wurden unverdünnt verwendet:
- PE Mouse Anti-Human CD42a, BD PharmingenTM
- FITC PAC-1, BD PharmingenTM
- PE-CyTM5 Mouse Anti-Human CD62P, BD PharmingenTM
- Arg-Gly-Asp-Ser (RGDS), Sigma
- PE-Cy™5 Mouse IgG1 κ Isotype Control, BD PharmingenTM
Anschließend wurde jede Probe mit 70 µl Cellfix (BD Biosciences) für 10 Min.
bei RT fixiert und danach mit 1 ml DPBS (mit Ca2+, Mg2+, 0,2% BSA)
verdünnt. Die Messung erfolgte innerhalb von 4 h am Durchflusszytometer
FC500 der Firma Beckmann-Coulter unter Verwendung eines geeigneten
FACS-Messprotokolls.
3.6.2 Durchflusszytometrie muriner Zellen
Die Überprüfung der Reinheit der isolierten CD4+-Zellpopulationen aus der
Milz erfolgte mit dem Durchflusszytometer.
Folgende Antikörper wurden unverdünnt verwendet:
- PE conjugated anti-mouse CD45RB (120455), eBioscience
- FITC conjugated monoclonal anti-mouse CD4 (130-091-608),
Miltenyi Biotec
Page 43
Kapitel 3: Material und Methoden
35
Nach der magnetischen Separation wurden 1x106 Zellen in 100 µl FACS-
Puffer aufgenommen und mit 3 µl Fc-Block 10 Min. inkubiert. Pro Probe
wurden je 1 µl CD45RB- und 3 µl CD4-Antikörper dazugegeben und 20 Min.
inkubiert. Nach einmaligem Waschen mit 2 ml FACS-Puffer (versetzt mit 2%
FCS) und 10 Min. Zentrifugation bei 500xg wurden die gefärbten Zellen in
250 µl FACS-Puffer aufgenommen und durchflusszytometrisch analysiert.
3.6.3 Fibrinogen-ELISA
Der Fibrinogen-ELISA funktioniert nach dem Prinzip eines Sandwich-ELISAs.
Das Fibrinogen im murinen Plasma bindet an die mit anti-Fibrinogen
beschichtete Oberfläche der 96-Well-Mikrotiterplatte. Plasmaproben wurden
vorher aus antikoaguliertem Mausblut mittels 10 Min. Zentrifugation bei
13000xg gewonnen und vor Auftragen 1:30000 mit Diluent-Konzentrat
verdünnt. Anschließend wurde der ELISA entsprechend der
Herstellerangaben durchgeführt. Die Absorption wurde bei 450 nm gemessen.
Mithilfe einer Standardkurve konnte die absolute Fibrinogen-Konzentration im
Plasma bestimmt werden.
3.6.4 Immunhistologie
3.6.4.1 Anfertigen von histologischen Präparaten
Die Anfertigung der Kryoschnitte erfolgte im Mikrotom-Kryostat bei -20°C
Kammertemperatur. Jeweils zwei 5 µm dicke Schnitte wurden auf Superfrost
Objektträger gebracht und ü.N. bei RT luftgetrocknet. Zur Fixation wurden die
Schnitte für 10 Min. in eiskaltes Aceton getaucht und anschließend eine
Stunde an der Luft getrocknet. Bis zur Färbung wurden sie bei -80°C gelagert.
3.6.4.2 Färbungen
Von Leber, Lunge, Milz und Niere wurden immunhistochemische Färbungen
angefertigt. Die Analyse der gefärbten Präparate erfolgte mit Hilfe des Leica
DM IL Mikroskops (Leica Microsystems, Wetzlar). Fotografiert wurden die
Schnitte mit einer Kamera von Visitron System GmbH und der zugehörigen
Software Spot Advanced (Diagnostic Instruments Inc.) und eingefärbt mit
MetaMorph Offline. Ausgewertet wurden die Bilder anschließend mit der
Software ImageJ.
Page 44
Kapitel 3: Material und Methoden
36
Immunhistochemische Färbungen
Zur immunhistochemischen Färbung der Organe wurden spezifische
monoklonale Antikörper verwendet, die an die nachzuweisenden
Oberflächenepitope binden. Zur Sichtbarmachung der Bindung wurden mit
einem Fluoreszensfarbstoff gekoppelte Antikörper, die Peroxidase-
Antiperoxidasemethode, sowie die Avidin-Biotin-Methode verwendet.
Folgende Primärantikörper wurden verwendet:
- FITC-labeled rat monoclonal anti-mouse GPIbβ (CD42c) (M050-1), emfret
analytics Eibelstadt
- Purified Rat anti-mouse Ly-6G and Ly-6c (553123), BD PharmingenTM,
Heidelberg
- Purified rat anti-mouse CD62P (550289), BD PharmingenTM, Heidelberg
Folgende Sekundär- und Tertiärantikörper wurden verwendet:
- POD conjugated F(ab’)2 Fragment mouse anti-ratIgG (H+L), (212-036-
168), Jackson Immuno Research
- Alexa 488 (A-11090), Molecular Probes
- Streptavidin TRITC (016-030-084), Dianova
- TSATM Biotin System, für 200-600 Proben (NEL700001KT), PerkinElmer
Die Färbungen wurden wie folgt durchgeführt:
Kombinationsfärbung von Thrombozyten mit P-Selektin oder GR-1
Der gesamte Färbevorgang fand in einer feuchten Kammer statt um die
Austrocknung der Präparate zu verhindern. Waschschritte erfolgten jeweils für
4 Min. in PBS. Zuerst wurden die Organe mit 20 µl 20%igem fetalem
Kälberserum in PBS 10 Min. inkubiert um unspezifische Fc-Bindungsstellen zu
blocken. Anschließend wurden mittels Peroxidase-Blocking-Reagenz (10 Min.,
Ready-to-use, Dako, Carpinteria, USA) die endogenen Peroxidasen inaktiviert
um eine unerwünschte Aktivierung der später zugegebenen chromogenen
Page 45
Kapitel 3: Material und Methoden
37
Substanzen zu verhindern. Nach zweimaligem Waschen erfolgte die Zugabe
eines Tropfens Biotin-Blocking-Lösung 1 (Avidin Solution, Dako X0590) für 10
Min. um eine Bindung des enzymmarkierten (TRITC) Streptavidins an
endogenem Biotin zu verhindern. Damit werden falsch-positive Ergebnisse
vermieden. Es wurde einmal gewaschen um dann Biotin-Bindungsstellen
mittels einem Tropfen Biotin-Blocking-Lösung 2 (Biotin Solution, 10 Min.) zu
blockieren. Nach nochmaligem Waschen erfolgte die Zugabe von 100 µl des
primären Antikörpergemisches (CD62P, 1:50 verdünnt in 20% FCS/PBS und
CD 42c FITC 1:20 verdünnt in 20% FCS/PBS bzw. CD 42c FITC 1:20
verdünnt in 20% FCS/PBS und Gr-1 1:50 in PBS/ 20% FCS) bzw. von 100 µl
20% FCS/PBS auf die Negativkontrolle. Die Schnitte wurden ü.N. bei 4°C
inkubiert. Darauf wurde dreimal gewaschen und schließlich der Peroxidase
gekoppelte Sekundärantiköper zugegeben (anti rat IgG POD 1:100 in PBS
/20% FCS) und 60 Min. bei RT inkubiert. Anschließend wurden die Schnitte
dreimal gewaschen, mit 50 µl Biotinyltyramid (1:200 verdünnt in
Amplifikationsdiluent) versehen und 4 Min. inkubiert. Nach zweimaligen
Waschen wurden 50 µl des dritten Antikörpergemisches (Streptavidin TRITC
1:100 verdünnt in PBS/20%FCS, aFITC-Alexa 488 1:100 verdünnt in
PBS/20%FCS) dazugegeben und 60 Min. bei RT inkubiert. Nach zweimaligem
Waschen wurden die Schnitte mit Fluoprep eingedeckelt.
Einfachfärbung von Thrombozyten
Die Färbung erfolgte analog wie oben beschrieben. Im Unterschied zur
kombinierten Färbung wurde nach Auftragen von 75 µl des Primärantikörpers
(CD 42c FITC 1:20 verdünnt in 20% FCS/PBS, Inkubation über Nacht bei 4°C)
auf die Positivkontrolle nur zweimal gewaschen, bevor der zweite Antikörper
zur Verstärkung des Signals dazugegeben wurde. Nachdem 50 µl des
Verstärkungsantikörpers Alexa 488 (1:100 verdünnt in PBS/20%FCS)
dazugegeben wurden, wurden die Schnitte zweimal gewaschen und mit
Fluoprep eingedeckelt.
Page 46
Kapitel 3: Material und Methoden
38
3.7 Statistische Auswertung
Die statistische Auswertung der Ergebnisse und die Darstellung der Graphen
erfolgte mit dem Programm GraphPad Prism für Windows (GraphPad
Software, San Diego, CA).
Jedes Ereignis entspricht einer Probe bzw. einem Tier. Dargestellt ist der
arithmetische Mittelwert. Die Daten wurden mittels des Mann-Whitney-Test auf
Signifikanz überprüft, da eine Normalverteilung der Daten nicht gewährleistet
werden kann. Als Signifikanzniveau wurde p = 0,05 festgelegt und in den
Graphen mit * verdeutlicht. Zwei ** stehen für ein Signifikanzniveau von
p< 0,01 und drei *** für p< 0,001.
Die Auswertung der FACS Analysen erfolgte mit FlowJo. Die Gates wurden
entsprechend der Größe (Vorwärtsscatter) und Granularität (Seitwärtsscatter)
der Zellen gesetzt um anschließend die populationsspezifische Analyse der
Bindung der Farbstoff-gekoppelten Antikörper durchzuführen. Histogramme
und Dotplots wurden mit Hilfe von FlowJo dargestellt und ausgewertet.
Page 47
Kapitel 4: Ergebnisse
39
4 Ergebnisse
4.1 In vitro Versuche mit humanen Zellen
Um die Rolle der Thrombozyten in der polymikrobiellen Sepsis zu
untersuchen, wurden in vitro Versuche mit humanen Zellen durchgeführt. Als
vereinfachtes Sepsismodell diente hierbei die Stimulation von Zellen mit LPS
(E.coli O127:B8). Überprüft wurde, ob Thrombozyten durch diesen
Bakterienbestandteil aktiviert werden und welche Bedingungen dafür nötig
sind.
4.1.1 Stimulation von plättchenreichem Plasma
Es wurden sowohl Versuche mit LPS als auch Kostimulationsversuche mit
LPS und TRAP-6, einem synthetischen Thrombinanalogon, durchgeführt um
zu untersuchen, ob LPS selbst Thrombozyten aktiviert oder eine Verstärkung
der Aktivierung durch TRAP-6 auslöst.
Thrombozyten weisen in der Durchflusszytometrie eine charakteristische
Population auf, die sich bei Maximalaktivierung in ihrer Form ändert. Die
Änderung der Thrombozytenwolke ist bedingt durch den „shape change“ des
Thrombozyts nach Aktivierung mit Thrombin bzw. TRAP-6 [38].
Abb. 3: Typische Thrombozytenpopulation in der Durchflusszytometrie
Dargestellt ist eine charakteristische Thrombozytenwolke im Vorwärts-Seitwärtsscatter des
Durchflusszytometers. Der Übergang der diskoiden, nicht-aktivierten Thrombozyten (links) zu
sphärischen Partikeln nach Maximalaktivierung mit 200 µmol/l TRAP-6 (rechts) wird auch
durchflusszytometrisch sichtbar.
Page 48
Kapitel 4: Ergebnisse
40
Da die Formänderung der Thrombozytenpopulation nur bei sehr starker
Aktivierung beobachtet werden kann, wurden zusätzlich farbstoffmarkierte
Aktivierungsmarker verwendet um kleinere Unterschiede in der Aktivierung
festzustellen.
Als Fluoreszenzmarker wurden die Antikörper der Aktivierungsmarker
P-Selektin (CD62P) und PAC-1 verwendet und deren Bindung
durchflusszytometrisch untersucht. P-Selektin wird nach Aktivierung der
Thrombozyten aus den α-Granula freigesetzt und als Membranprotein auf der
Oberfläche exprimiert. PAC-1 bindet an den aktivierten Fibrinogenrezeptor,
der nach entsprechender Stimulation durch Formkonversion des
Oberflächenepitops GPIIb/IIIa zu einem funktionellen Rezeptor für Fibrinogen
entsteht. Besonders PAC-1 ist ein sehr sensitiver Aktivierungsmarker, der
schon bei leichter Stimulation der Plättchen exprimiert wird.
4.1.1.1 Stimulation mit TRAP-6
Vor Beginn der Versuche wurde ein Titrationsversuch mit TRAP-6
durchgeführt um die optimale Konzentration für eine Kombination mit LPS zu
finden.
Abb. 4: CD62P-Expression nach Stimulation von PRP mit unterschiedlichen TRAP-6
Konzentrationen
Page 49
Kapitel 4: Ergebnisse
41
5x106 Thrombozyten wurden mit 200 µmol/l, 50 µmol/l, 12,5 µmol/l TRAP-6 und als Kontrolle
mit PBS stimuliert und anschließend die P-Selektin-Expression durchflusszytometrisch
bestimmt. 200 µmol/l TRAP-6 aktivierte beinah alle Thrombozyten (dunkelrot links, hellgrau
rechts), während mit 12,5 µmol/l TRAP-6 (rosa) kein Unterschied zur Kontrollgruppe
(dunkelgrau) zu erkennen war. Durch Zugabe von 50 µmol/l TRAP-6 kam es zu einer mäßigen
Aktivierung der Thrombozyten (rot).
In Abb. 4 ist sowohl im Histogramm als auch im Dotplot ersichtlich, dass es
nach Zugabe von 200 µmol/ TRAP-6 zu einer maximalen Degranulation der
α-Granula und Expression von CD62P und damit zu einer starken Aktivierung
kam. Bei zusätzlicher Behandlung mit LPS wäre eine Steigerung der
Aktivierung kaum noch möglich, da gespeichertes P-Selektin in den α-Granula
bereits verbraucht wurde. Im Gegensatz dazu war nach Stimulation mit
12,5 µmol/l TRAP-6 kein Unterschied zur PBS-behandelten Kontrollgruppe zu
erkennen. Die Zellen, die mit 50 µmol/l TRAP-6 stimuliert wurden, zeigten eine
mäßige Steigerung der P-Selektin-Expression. Dies ist zu erkennen an der im
Histogramm dargestellten mittleren Fluoreszenz und an der Anzahl CD62P-
positiver Ereignisse im Dotplot. Da hier eine optimale Beobachtung der
Änderung einer Expressionsrate und Aktivierung möglich ist, wurde diese
Konzentration zur Kostimulation mit LPS verwendet.
4.1.1.2 Stimulation mit TRAP-6 und LPS
Blutproben wurden von freiwilligen, gesunden Spendern gewonnen und mit
Citrat antikoaguliert. Jeweils 5x106 Zellen des hergestellten PRP wurden pro
Röhrchen überführt und mit LPS und TRAP-6 stimuliert (siehe Tabelle 1).
Pro Gruppe wurden Proben von sieben Spendern untersucht. Der
Versuchsaufbau ist in Tabelle 1 dargestellt.
Page 50
Kapitel 4: Ergebnisse
42
Tabelle 1: Versuchsaufbau der Stimulation von PRP
Nach Inkubation der Stimulanzien wurden die Aktivierungsmarker CD62P und
PAC-1 dazugegeben und deren Expression durchflusszytometrisch bestimmt.
Sample TRAP-6
(50 µmol/l)
LPS
O127:B8
(10 µg/ml)
RGDS
10 mg/ml
(je 5 µl)
PAC-1
FITC
Unv.
(je 5 µl)
CD62P
PE-Cy5
Unv.
(je 5 µl)
IgG1
PE-Cy5
Unv.
(je 5 µl)
1 + - - + + -
2 + - + + - +
3 + + - + + -
4 + + + + - +
5 - - - + + -
6 - - + + - +
7 - + - + + -
8 - + + + - +
Page 51
Kapitel 4: Ergebnisse
43
Abb. 5: Histogramme der CD62P- und PAC-1-Fluoreszenz nach Stimulation mit TRAP-6
und LPS
Gezeigt ist die Fluoreszenzintensität der Aktivierungsmarker CD62P und PAC-1. In den
Histogrammen links erfolgte die Zugabe von PBS als Kontrolle (grau) und die zusätzliche
Stimulation mit 10 µg/ml LPS (rot). Die rechten Graphen zeigen die Fluoreszenz der
Aktivierungsmarker einmal nach Zugabe von 50 µmol/l TRAP-6 als Kontrolle (grau) und
zusätzlicher Stimulation mit LPS (10 µg/ml) (rot). Die Fluoreszenz nach LPS-Zugabe steigt bei
beiden Gruppen nur minimal an.
Wie in Abb. 5 zu erkennen ist, steigt die Fluoreszenz der Aktivierungsmarker
nach LPS-Zugabe nur minimal, es erfolgt also keine merkliche Stimulation
durch LPS. Auch eine signifikante Steigerung der Aktivierung von TRAP-6
durch LPS konnte nicht beobachtet werden.
Page 52
Kapitel 4: Ergebnisse
44
TRAP-6 TRAP-6+LPS Ko LPS0
500
1000
1500
2000
PAC-1
CD62PG
eom
etr
ic M
ean
Abb. 6: Mittlere Fluoreszenz von PAC-1 und P-Selektin nach Stimulation
Dargestellt ist der Mittelwert der Fluoreszenz von PAC-1 (grau) und P-Selektin (rot). PRP
wurde entweder mit 50 µmol/l TRAP-6 behandelt oder als Kontrolle (Ko) mit PBS. Zusätzlich
wurde jeweils mit 10 µg/ml LPS (E.coli) stimuliert. Die Expression von P-Selektin ist nach
TRAP-6-Stimulation signifikant höher, als in der Kontroll- und LPS-Gruppe (p<0,05). Durch
LPS-Stimulation erhöht sich die mittlere Fluoreszenz nur geringfügig (n=7 pro Gruppe).
Thrombozyten im plättchenreichen Plasma zeigen nach Stimulation mit LPS
keine signifikante Erhöhung der Expression vom Fibrinogen-Rezeptor (p=0,65)
und P-Selektin (p=0,41) im Vergleich zur PBS behandelten Kontrollgruppe,
werden also nicht aktiviert. Auch eine signifikante Steigerung der Aktivierung
durch LPS nach Kostimulation mit TRAP-6 konnte sowohl bei Pac-1 (p=0,58)
als auch bei P-Selektin (p=0,59) nicht beobachtet werden, die Aktivierung von
TRAP-6 konnte also durch LPS nicht verstärkt werden. Zu sehen sind nur
geringe Anstiege in der Fluoreszenz der Aktivierungsmarker (Abb. 6).
Thrombozyten werden also im PRP nicht durch LPS aktiviert.
4.1.2 Stimulation von Vollblut
Um zu überprüfen ob Thrombozyten im Vollblut, also in Kombination mit
anderen Blutzellen, durch LPS aktiviert werden, wurden jeweils 100 µl Vollblut
von freiwilligen Spendern mit LPS, TRAP-6 oder beidem stimuliert. Als
Kontrolle wurde PBS hinzugegeben. Nach zwei Stunden Inkubation wurden
ein Antikörper zur Messung der Thrombozytenaktivierung (CD62P) und ein
thrombozytenspezifischer Antikörper (CD41) zur Bestimmung der Aggregate
mit anderen Blutzellen hinzugegeben.
Der Versuchsaufbau ist in Tabelle 2 dargestellt.
Page 53
Kapitel 4: Ergebnisse
45
Tabelle 2: Versuchsaufbau der Stimulation von Vollblut
Sample TRAP-6
50µmol/l
PBS
(+Ca/+Mg)
LPS O127:B8
10µg/ml
Antikörper
(je 5µl)
1 - + - Isotyp Tricolor
2 - + - CD45 FITC,
CD41 PC5
3 - + - CD45 FITC,
CD62P PERCP
4 - + + Isotyp Tricolor
5 - + + CD45 FITC,
CD41 PC5
6 - + + CD45 FITC,
CD62P PERCP
7 + - - Isotyp Tricolor
8 + - - CD45 FITC,
CD41 PC5
9 + - - CD45 FITC,
CD62P PERCP
10 + - + Isotyp Tricolor
11 + - + CD45 FITC,
CD41 PC5
12 + - + CD45 FITC,
CD62P PERCP
P-Selektin diente als Aktivierungsmarker, da es nur auf aktivierten
Thrombozyten und Endothelzellen exprimiert wird [39]. Als spezifischer Marker
für Thrombozyten wurde CD41, Teil des Rezeptors GPIIb/IIIa, verwendet. Die
verschiedenen Populationen im Vollblut wurden über den Vorwärts-Seitwärts-
Scatter differenziert (Abb. 7)
Page 54
Kapitel 4: Ergebnisse
46
Abb. 7: Durchflusszytometrische Darstellung der Zellen im Vollblut
Die Gates wurden entsprechend der zellspezifischen Morphologie Größe und Granularität
gesetzt. Die festgelegten Zellpopulationen Monozyten, Granulozyten und Lymphozyten
wurden anschließend entsprechend ihrer Gates näher analysiert. Ein Punkt entspricht einem
Ereignis im FACS.
4.1.2.1 Granulozyten
Zuerst wurden die Aggregate der Thrombozyten und Granulozyten untersucht,
also die CD41-positiven Zellen in der vorher morphologisch bestimmten
Granulozytenpopulation. Alle untersuchten Granulozyten waren CD45R
positiv.
Abb. 8: Zunahme der Thrombozyten-Granulozyten-Aggregate
Page 55
Kapitel 4: Ergebnisse
47
CD
41
In der unbehandelten Probe waren kaum CD41-positive Granulozyten zu sehen. Nach LPS-
Stimulation stieg das CD41-Signal deutlich an. Der Anteil an Aggregaten lag in der
unbehandelten Gruppe bei 17,2% und stieg durch LPS Stimulation auf 39,49%. 100 µl Vollblut
wurden mit LPS stimuliert (E.coli, 10 µg/ml) und 2 h inkubiert. Durchflusszytometrische Daten
wurden mit FlowJo ausgewertet. Die Gates wurden mithilfe einer Isotypkontrolle festgelegt.
Nach LPS-Stimulation steigt das Fluoreszenzsignal von CD41 in der
Granulozytenpopulation deutlich an. Es kommt also zur Erhöhung der
Thrombozyten-Granulozyten-Aggregate. Eine Erhöhung der Zell-Zell-
Aggregate ließ sich auch durch TRAP-6-Zugabe erzeugen. Durch
Kostimulation von TRAP-6 mit LPS konnte ein zusätzlicher Anstieg der CD41-
positiven Zellen beobachtet werden (Abb. 9).
PBSLPS
TRAP-6
TRAP-6 + LPS
0
20
40
60
80
100**
**
CD
41 in
%
Abb. 9: Unterschiedliche Expression des Thrombozytenmarkers CD41 in der
Granulozytenpopulation nach Stimulation mit LPS und TRAP-6
Nach Simulation von Vollblut mit LPS war die CD41-Expression auf den Granulozyten
signifikant höher als in der Kontrollgruppe (PBS) (p<0,01). Durch Zugabe des
Thrombozytenaktivators TRAP-6 stieg die CD41-Fluoreszenz im Vergleich zur PBS-Gruppe
auch signifikant an (p<0,01). Die Kostimulation von TRAP-6 mit LPS erhöht zusätzlich die
Aggregatbildung zwischen Granulozyten und Thrombozyten (p=0.0503).
Neben der Aggregatbildung wurde auch die Aktivierung der Thrombozyten, die
sich in diesen Aggregaten befanden, untersucht. Hierzu wurde die Expression
von P-Selektin mithilfe eines anti-CD62P-Antkörpers untersucht.
Page 56
Kapitel 4: Ergebnisse
48
Abb. 10: CD62P -Expression der Thrombozyten in der Granulozytenpopulation
Die LPS-behandelte Gruppe (rot) zeigt im Vergleich zur PBS-behandelten Kontrollgruppe
(grau) eine deutliche Zunahme der Fluoreszenz von CD62P (links). Die rechte Graphik zeigt,
dass LPS als Kostimulanz mit TRAP-6 (rot) zusätzlich die Fluoreszenz des
Aktivierungsmarkers CD62P auf den Thrombozyten in der Granulozytenpopulation erhöht im
Vergleich zur TRAP-6 behandelten Kontrollgruppe (grau). Die Isotypkontrolle ist in den
Histogrammen gestrichelt dargestellt. 100 µl Vollblut wurden mit LPS (10 µg/ml) bzw. TRAP-6
(50 µmol/l) oder beidem stimuliert.
Die Expression von P-Selektin steigt nach Zugabe von LPS in jeder Gruppe
signifikant an. (Abb. 11)
PBSLPS
TRAP-6
TRAP-6 + LPS
0
10
20
30
40
50
60
70 *** *
**
CD
62P
in %
Abb. 11: Unterschiedliche Aktivierung der Thrombozyten in der
Granulozytenpopulation
Page 57
Kapitel 4: Ergebnisse
49
Aktivierung wurde durch P-Selektin-Expression der Thrombozyten bestimmt. Nach LPS-
Stimulation stieg die Aktivierung der Thrombozyten signifikant an im Vergleich zur PBS-
behandelten Kontrollgruppe (p<0,001). Auch durch Kostimulation war ein signifikanter Anstieg
der P-Selektin-Fluoreszenz zu beobachten im Vergleich zur TRAP-6 behandelten Gruppe
(p<0,05). 100µl Vollblut wurden mit LPS (10µg/ml) bzw. TRAP-6 (50µmol/l) stimuliert und 4 h
inkubiert (n=9 pro Gruppe).
4.1.2.2 Monozyten
Zusätzlich zur Population der Granulozyten wurde die Monozytenpopulation
näher betrachtet und auf dieselben Parameter wie bei den Granulozyten
analysiert.
Abb. 12: Zunahme der Thrombozyten-Monozyten-Aggregate nach LPS-Stimulation
Vollblut wurde mit LPS (10 µg/ml) stimuliert und 4 h inkubiert. CD41-Expression wurde
durchflusszytometrisch bestimmt. Zu sehen war eine deutliche Erhöhung der CD41-
Fluoreszenz in der Monozytenpopulation von 50,94% in der unbehandelten auf 68,93% in der
stimulierten Gruppe. Daten wurden ausgewertet mit FlowJo. Zur Festlegung der Gates diente
die Isotypkontrolle.
Die CD41-Expression der Monozyten nahm nach LPS-Zugabe signifikant zu
(p<0.01). Durch LPS banden Thrombozyten vermehrt an die Monozyten im
Vollblut. Durch TRAP-6 wurde ein ähnlicher Effekt, wie durch LPS ausgelöst.
Nach zusätzlicher Stimulation von LPS in Kombination mit TRAP-6 stieg die
CD41-Fluoreszenz nur noch geringfügig an. (Abb. 13)
Page 58
Kapitel 4: Ergebnisse
50
PBSLPS
TRAP-6
TRAP-6 + LPS
0
20
40
60
80
100**
**
CD
41+
in %
Abb. 13: CD41-Expression in der Monozyten-Population nach LPS und TRAP-6 Zugabe
CD41-Fluoreszenz wurde durchflusszytometrisch im Vollblut gemessen zur Bestimmung der
Monozyten-Thrombozyten-Aggregate. In der Monozytenpopulation stieg der Anteil an CD41-
positiven Zellen signifikant an nach LPS- und TRAP-6-Stimulation (p<0.01). Eine Kombination
von LPS mit TRAP-6 zeigte keine Erhöhung der CD41-Expression im Vergleich zur einfachen
TRAP-6-Behandlung. 100 µl Vollblut wurden mit LPS (10 µg/ml) bzw. TRAP-6 (50 µmol/l)
stimuliert und 4 h inkubiert (n=8-10 pro Gruppe).
Wie für die Granulozyten gezeigt, nimmt auch bei den Monozyten die CD62P-
Expression zu. Thrombozyten, die an die Monozyten gebunden haben, zeigen
nach Stimulation mit LPS eine stärkere Aktivierung. Die Zunahme der
Fluoreszenzintensität von CD62P ist in den Histogrammen zu erkennen. (Abb.
14)
Abb. 14: CD62P-Expression der Thrombozyten in der Monozytenpopulation
Page 59
Kapitel 4: Ergebnisse
51
Nach alleiniger Zugabe von LPS (rot) zum Vollblut oder als Kostimulanz mit TRAP-6 erhöhte
sich die Fluoreszenz des Aktivierungsmarkers CD62P auf den Thrombozyten in der
Monozytenpopulation im Vergleich zur jeweiligen Kontrollgruppe (grau). Betrachtet wurde die
gesamte Monozytenpopulation. Die Isotypkontrolle ist gestrichelt dargestellt.
Bei Betrachtung aller Gruppen fällt eine starke Aktivierung der Thrombozyten
nach LPS-Zugabe auf, sowohl im Vergleich zur PBS-behandelten
Kontrollgruppe als auch zur TRAP-6-behandelten Gruppe. Die Stimulation mit
LPS zeigt tendenziell sogar einen stärkeren Anstieg von CD62P als die
alleinige Stimulation mit TRAP-6. Die Unterschiede zwischen den einzelnen
Gruppen weisen Signifikanzen auf (Abb. 15).
PBS
LPS
TRAP-6
TRAP-6
+ L
PS
0
20
40
60
80 *** ****
CD
62P
in %
Abb. 15: Unterschiedliche Aktivierung der Thrombozyten in der Monozytenpopulation
Die Aktivierung wurde durchflusszytometrisch bestimmt mittels der P-Selektin-Expression der
Thrombozyten. Nach LPS-Stimulation stieg die Fluoreszenz von P-Selektin auf den
Thrombozyten signifikant an im Vergleich zur PBS-behandelten Kontrollgruppe (p<0,001).
Auch durch Kostimulation von TRAP-6 und LPS ist ein signifikanter Anstieg der P-Selektin-
Fluoreszenz zu beobachten im Vergleich zur TRAP-6 behandelten Gruppe (p<0,05).
Im Vollblut kam es also zu einer Aktivierung von Thrombozyten durch LPS.
Die Aktivierung äußerte sich über eine Zunahme der P-Selektin-Expression
und über eine vermehrte Bildung von Zell-Zell-Aggregaten zwischen
Thrombozyten und Granulozyten bzw. Monozyten.
Page 60
Kapitel 4: Ergebnisse
52
4.2 Versuche mit murinen Thrombozyten
Um das Verhalten von Thrombozyten während einer Sepsis in vivo zu
untersuchen, wurden murine Zellen von unbehandelten und CASP-operierten
Tieren verglichen. Die Versuche fanden jeweils 20 Stunden nach der
Operation statt.
4.2.1 Thrombozytenzahl nach CASP
Ein bekanntes Phänomen bei septischen Patienten ist die Thrombozytopenie.
Um die Übertragbarkeit auf das Tiermodell der CASP zu überprüfen, wurden
die Thrombozytenzahlen von unbehandelten und septischen Tieren verglichen
(Abb. 16).
WT CASP0
50
100
150
200
250
**
Thrm
obozyt
enza
hl x10
6/m
l
Abb. 16: Änderung der Thrombozytenzahl nach CASP
20 h nach CASP kam es zu einem signifikanten Abfall der Thrombozyten im Vollblut im
Vergleich zu unbehandelten Kontrolltieren (p<0.01). Das Blut wurde retroorbital entnommen.
Anschließend wurden die Plättchen isoliert und gezählt.
Die Thrombozytenzahl unbehandelter Tiere betrug nach Isolation der Zellen
im Mittel 161x106/ml. Nach CASP kam es zu einem signifikanten Abfall
(p<0.01) der Thrombozyten auf durchschnittlich 107x106/ml Zellen. Untersucht
wurden jeweils zehn Tiere.
CASP löste 20 h nach Operation bei den Tieren eine Thrombozytopenie aus.
4.2.2 Immunhistologischer Vergleich septischer und unbehandelter Organe
Um die Verteilung der Thrombozyten nach der CASP zu untersuchen und das
Verschwinden der Plättchen nachzuvollziehen, wurden immunhistologische
Page 61
Kapitel 4: Ergebnisse
53
Färbungen von Leber, Lunge, Milz und Niere angefertigt. Die Blutplättchen
wurden mit dem spezifischen Marker CD42c auf den Organschnitten
angefärbt. CD42c wird auch bezeichnet als GpIbβ und ist Teil des Komplexes
GpIa-d, der als Bindungsstelle für den von Willebrand-Faktor (vWF) und
Thrombin dient. Erhöht sich die mittlere Fluoreszenz von CD42c, lässt sich
daraus eine Erhöhung der Thrombozytenzahl in dem gefärbten Organ
herleiten. Verglichen wurden unbehandelte CCR4-/- und C57BL/6 Tiere mit
septischen 20 h nach CASP.
4.2.2.1 Ergebnisse von Lunge und Milz in CCR4-/- und C57BL/6
In Lunge und Milz zeigte sich kein Unterschied in der Anzahl angefärbter
Thrombozyten, sowohl zwischen den unbehandelten Tieren der C57BL/6 und
CCR4-/- Maus, als auch zwischen der CASP-Gruppe und der unbehandelten
Kontrollgruppe (Abb. 17).
Page 62
Kapitel 4: Ergebnisse
54
Abb. 17: Immunhistologische Färbungen von Lunge und Milz
Die Organe wurden unbehandelt oder 20 h nach CASP von C57BL/6 oder CCR4-/-
-Tieren
entnommen und mit einem FITC-markierten CD42c-Antikörper angefärbt um die
Thrombozyten sichtbar zu machen. Die Bilder zeigen die Fluoreszenz der gebundenen
Antikörper bei 100facher Vergrößerung.
20 h nach CASP kam es zu keiner Einwanderung von Thrombozyten in die
Lunge und die Milz. Die Organe konnten also nicht mit der Thrombozytopenie
in Zusammenhang gebracht werden (Abb. 18, Abb. 19).
Page 63
Kapitel 4: Ergebnisse
55
WT C
57Bl6
CASP C57Bl6 -/-
WT C
CR4-/-
CASP CCR4
0
1
2
3
4
Thro
mbozyte
nfläche in %
Abb. 18: Änderung der Thrombozytenzahl in der Lunge
CASP führte zu keiner Änderung der Thrombozytenzahlen in der Lunge, weder in C57BL/6
noch in CCR4-/-
Mäusen. Pro Gruppe erhielten 4 Mäuse CASP, 5 dienten als unbehandelte
Kontrolle. 20 h nach CASP wurden die Organe entnommen und mit dem Thrombozyten-
spezifischen Marker CD42c gefärbt. Die fotografierten Färbungen wurden mit ImageJ
ausgewertet.
WT C57Bl6
CASP C57Bl6 -/-
WT CCR4
-/-
CASP CCR4
20
30
40
50
Thro
mbozyte
nfläche in %
Abb. 19: Änderung der Thrombozytenzahl in der Milz
Nach CASP kam es zu keiner Änderung der Thrombozytenzahlen in der Milz, weder in
C57BL/6 noch in CCR4-/-
Mäusen. Pro Gruppe erhielten vier Mäuse CASP, fünf dienten als
unbehandelte Kontrolle. 20 h nach CASP wurden die Organe entnommen und mit dem
Thrombozyten-spezifischen Marker CD42c gefärbt. Die fotografierten Färbungen wurden mit
ImageJ ausgewertet.
Page 64
Kapitel 4: Ergebnisse
56
4.2.2.2 Ergebnisse von Leber
In der Leber konnte 20 h nach CASP-Induktion ein Anstieg der CD42c-
Fluoreszenz in den immunhistologischen Färbungen beobachtet werden. Es
kam also zu einer signifikanten Erhöhung der Thrombozytenzahl in der Leber
der septischen Tiere. Dieser Anstieg war sowohl in den CASP-behandelten
C57BL/6 (p<0,0001) als auch in den CCR4-/--Tieren (p=0,0159) zu sehen. Als
zweiter Zeitpunkt wurde der 48 h nach CASP Zeitpunkt für die Entnahme der
Lebern gewählt. Die Thrombozytenzahl in der Leber der C57BL/6-Tiere waren
zu diesem Zeitpunkt immer noch signifikant höher als in der Kontrollgruppe
(p=0,029) (Abb. 20, Abb. 21)
Abb. 20: Immunhistologische Färbungen der Leber
Page 65
Kapitel 4: Ergebnisse
57
Die Leber wurde 20 h nach CASP mit dem thrombozytenspezifischen Antikörper CD42c
gefärbt. Untersucht wurden jeweils CCR4-/-
- und C57BL/6 Tiere. Unbehandelte Tiere dienten
als Kontrolle. Die septischen Tiere zeigten eine Zunahme der Fluoreszenz im Vergleich zu
den Kontrolltieren (WT). Die Aufnahmen sind dargestellt in 100facher bzw. 200facher
Vergrößerung.
Abb. 21: Änderung der Thrombozytenzahl in der Leber
Die Leber wurde 20 h und 48 h nach CASP entnommen und mit dem Thrombozytenmarker
CD42c angefärbt. Verglichen wurden jeweils septische CCR4-/-
und C57BL/6 Tiere mit
unbehandelten Kontrollgruppen. Bei beiden Gruppen ist 20 h nach CASP ein signifikanter
Anstieg der Thrombozyten in den CASP-Tieren zu sehen (p<0.001 bei C57BL/6, links, p<0.05
bei CCR4-/-
, rechts). 48 h nach CASP nimmt die Anzahl der Thrombozyten in den C57BL/6-
Mäusen ab, ist aber noch signifikant erhöht im Vergleich zur Kontrollgruppe (p<0.01). Die
Bilder wurden ausgewertet mit ImageJ. Die Abbildung links fasst die Daten von fünf
unabhängigen Experimenten mit vergleichbaren Ergebnissen zusammen.
In den septischen Tieren kam es zu einer Sequestration von Thrombozyten in
der Leber, was somit als eine mögliche Ursache für die Thrombozytopenie
gesehen werden kann.
4.2.2.3 Ergebnisse der Leber mit CD42c , CD62P und Gr-1
Da die Anzahl der Thrombozyten nach CASP in der Leber zunahm, wurden
weitere Färbungen zur Untersuchung der Aktivierung und Aggregatbildung der
Thrombozyten mit Granulozyten in dem Organ angestellt. Dazu wurden
Kombinationsfärbung mit dem thrombozytenspezifischen Antikörper CD42c
und CD62P (P-Selektin) bzw. Gr-1 angefertigt. Gr-1 ist ein Oberflächenprotein
WT CASP 20h CASP 48h0.0
0.5
1.0
1.5
2.0
2.5 *****
Thro
mbozy
tenfläche in %
WT CASP 20h0
2
4
6
8
10 *
Thro
mbozyt
enflä
che in
%
Page 66
Kapitel 4: Ergebnisse
58
auf myeloiden Vorläuferzellen. Die Expression nimmt mit Reifung der
Granulozyten zu und wird als granulozytenspezifischer Marker genutzt.
Da die CCR4-/--Tiere keinen Unterschied zu den C57BL/6-Tieren zeigten,
wurde nur die Leber von Letzteren untersucht, jeweils 20 h nach CASP.
Die P-Selektin-Expression der Leber nahm 20 h nach CASP signifikant zu
(p<0.001) (Abb. 23). Durch Übereinanderlegen der P-Selektin- und CD42c-
Färbung konnte man sehen, dass die Fluoreszenzen kongruent waren.
P-Selektin wurde also ausschließlich von Thrombozyten exprimiert (Abb. 22).
Abb. 22: Immunhistochemische Anfärbung von P-Selektin und CD42c auf Leber
P-Selektin ist rot dargestellt, CD42c leuchtet grün. Überlagern sich beide Fluoreszenzen,
erscheint dies als gelb. Aufnahmen erfolgten bei 200facher Vergrößerung. Lebern wurden
20 h nach CASP entnommen und gefärbt. Organe unbehandelter Tiere (WT) dienten als
Kontrolle.
WT CASP 0
1
2
3
4
5**
Flä
che v
on P
-Sele
ktin
in %
Abb. 23: Zunahme der P-Selektin-Expression in der Leber nach CASP
Page 67
Kapitel 4: Ergebnisse
59
Lebern von C57BL/6 Tieren wurden 20 h nach CASP entnommen und mit CD42c und CD62P
angefärbt. Bei den septischen Tieren kam es im Vergleich zu den unbehandelten Tieren zu
einem signifikanten Anstieg der P-Selektin-Expression (p<0.01). Bilder wurden ausgewertet
mit ImageJ.
Mit Zunahme der Thrombozyten, nahm auch die CD62P-Expression der
septischen Tiere signifikant zu. In die Leber eingewanderte Thrombozyten
waren also aktiviert.
Bei der Färbung von CD42c und Gr-1 kam es 20 h nach CASP zu einer
deutlichen Zunahme beider Fluoreszenzen im Vergleich zur unbehandelten
Kontrollgruppe. Da es zu einer Überlagerung beider Fluoreszenzen kam,
lagen die Zellen offensichtlich als Aggregate vor. Die Infiltration der Leber mit
beiden Zelltypen war im septischen Tier signifikant höher als in der
unbehandelten Kontrollgruppe (p<0.01). Dargestellt in Abb. 25 ist nur die
Erhöhung der Granulozyten.
Abb. 24: Immunhistochemische Anfärbung von CD42c und Gr-1 in der Leber
20 h nach CASP war eine deutliche Zunahme beider Fluoreszenzen in der Leber zu erkennen.
Der Granulozytenmarker Gr-1 ist hier rot und CD42c als Thrombozytenmarker grün
dargestellt. Überlagerung beider Signale erscheint gelb. Die Aufnahmen wurden bei 200facher
Vergrößerung gemacht.
Page 68
Kapitel 4: Ergebnisse
60
WT CASP0
2
4
6
8**
Gra
nulo
zyt
enflä
che in
%
Abb. 25: Zunahme der Granulozyten in der Leber 20 h nach CASP
20 h nach CASP wurde die Leber entnommen und immunhistochemisch mit dem
Granulozytenmarker Gr-1 und dem Thrombozytenmarker CD42c gefärbt. Im Vergleich zu
unbehandelten Kontrolltieren stieg die Granulozytenzahl signifikant an (p<0.01). Bilder wurden
ausgewertet mit ImageJ.
Auch in vivo kam es nach CASP zu einer Zunahme von Thrombozyten-
Granulozyten-Aggregaten in Leber.
4.2.3 Fibrinogen-ELISA
Eine andere Ursache für das Verschwinden der Thrombozyten in der Sepsis
ist die disseminierte intravasale Gerinnung (DIC). Da es dabei neben der
Aggregation von Thrombozyten auch zu einer starken Aktivierung der
Gerinnungskaskade kommt, kann man einen Abfall von Fibrinogen im Plasma
beobachten. Bei der Gerinnung entsteht Thrombin, das zur Bildung von Fibrin
aus Fibrinogen führt. Es entstehen Thromben und Thrombozytenaggregate,
die sich in Gefäßen und Sinusoiden der Organe ablagern und zum
Organversagen führen können. Fibrinogen ist neben einem Marker für
schwere DIC auch ein akute-Phase-Protein und ist damit nach Traumen und
bei Entzündung erhöht.
Um zu untersuchen, ob es zu einem Abfall von Fibrinogen nach CASP kommt,
wurde ein Fibrinogen-ELISA durchgeführt. 20 h nach CASP wurde Plasma
von septischen Tieren (C57BL/6 und CCR4-/-, n=7-8 pro Gruppe) gewonnen
und 1:30000 verdünnt aufgetragen. Plasma von unbehandelten Kontrolltieren
(n=8-9) wurden parallel in der gleichen Verdünnung aufgetragen. Die
Page 69
Kapitel 4: Ergebnisse
61
Absorption wurde bei 450 nm gemessen und mittels einer Standardkurve
(Abb. 26) wurde die absolute Fibrinogenkonzentration im Plasma bestimmt.
0 200 400 600 800 10000
1
2
3
4
ng / ml
O.D
. 450 n
m
Abb. 26: Standardreihe für den Fibrinogen-ELISA
Angefertigte Standardreihe mit löslichem Fibrinogen. 800 ng/ml Fibrinogen entsprechen einer
Absorption von 3,2 nm. Erstellt mit GraphPadPrism.
Bei den septischen C57BL/6-Tieren kam es im Vergleich zur Kontrollgruppe
zu einem signifikanten Anstieg der absoluten Fibrinogenkonzentration im
Plasma von im Durchschnitt 1,94 mg/ml auf 5,17 mg/ml (p<0.001). Eine
Erhöhung des Fibrinogengehalts war auch bei den CCR4-/--Tieren zu sehen.
Hier stieg das Fibrinogen von im Mittel 1,97 mg/ml bei den Kontrolltieren auf
5,86 mg/ml bei den CASP-Tieren (p<0.001) (Abb. 27).
WT C
57BL/6
CASP C57BL/6
-/-
WT C
CR4-/-
CASP CCR4
0
5
10
15 ******
Fib
rinogen in m
g/m
l
Abb. 27: Zunahme von Fibrinogen im Plasma 20h nach CASP
Page 70
Kapitel 4: Ergebnisse
62
20 h nach CASP wurde Plasma gewonnen sowohl von C57BL/6 als auch von CCR4-/-
Mäusen. Der Fibrinogengehalt wurde mittels Sandwich-ELISA bestimmt. Im Vergleich zu den
unbehandelten Kontrolltieren kam es in beiden Gruppen zu einem signifikanten Anstieg von
Fibrinogen (p<0.001).
Trotz Vermutung kam es 20 h nach CASP nicht zu einem Abfall von
Fibrinogen im Plasma. DIC konnte nicht nachgewiesen werden.
4.3 CCR4-Regulation nach CASP
Da die CCR4-/--Mäuse bisher keinen Unterschied zu C57BL/6-Tieren
aufwiesen, nach CASP aber ein besseres Überleben zeigen [30], wurde die
Regulation von CCR4 auch in PBMCs und CD4+-Zellen untersucht. In einem
Parallelprojekt unserer Arbeitsgruppe konnte bereits gezeigt werden, dass
CCR4 auf mRNA-Ebene in Thrombozyten 20 h nach Sepsisinduktion
hochreguliert wird.
Die Transkriptionsrate von CCR4 und damit die Menge der CCR4-mRNA
wurde mittels relativer Echtzeit-PCR bestimmt. Als endogene Kontrolle diente
GAPDH. Verglichen wurde die RNA von CASP-Tieren mit der von
unbehandelten Kontrolltieren nach der ΔΔCt-Methode. Jeweils 20 h nach
CASP wurde Blut für die Isolation der PBMCs bzw. Milz für die Isolation von
CD4+-Zellen entnommen.
4.3.1 RT-qPCR muriner PBMCs
Murine PBMCs wurden 20 h nach CASP isoliert. Bei Zählung der Leukozyten
im Vollblut fiel auf, dass die Zahl der weißen Blutkörperchen insgesamt und
auch die der Lymphozyten signifikant abnahm im Vergleich zum
unbehandelten Kontrolltier. Leukozyten fielen von im Mittel 2,97 x109/ml auf
1,96 x109/ml. Der Anteil der Lymphozyten davon sank von 2,52 x109/ml auf
1,67 x 109/ml (Abb. 28).
Page 71
Kapitel 4: Ergebnisse
63
WT W
BZ
CAS
P W
BZ
WT L
YM
CAS
P L
YM
0
1
2
3
4
5* *
Zellz
ahl x
10
9/m
l
Abb. 28: Abnahme der Leukozyten 20 h nach CASP
Die Zahl der Leukozyten (WBZ) im Vollblut nahm 20 h nach CASP signifikant ab (p<0.01). Bei
Betrachtung der Lymphozytenpopulation (LYM) sinkt deren Zellzahl auch signifikant im
Vergleich zu unbehandelten Kontrolltieren (p<0.05). Vollblut wurde retrobulbär abgenommen
und die Zellzahl mit dem Sysmex Cellcounter bestimmt (n=6-7 pro Gruppe).
Die RNA wurde mit der Trizol-Methode isoliert und anschließend mit dem
NanoDrop auf ihre Qualität untersucht. Es wurde ausschließlich qualitativ
hochwertige RNA ohne Verunreinigung verwendet. Nach cDNA-Synthese
wurde die CCR4-Expression auf mRNA Ebene mit quantitativer Echtzeit-PCR
untersucht. Das house-keeping Gen GAPDH diente hierbei als endogene
Kontrolle. Verglichen wurde zunächst die Größe des Fluoreszenzsignals der
einzelnen farbstoffgekoppelten Primer (Abb. 29).
Page 72
Kapitel 4: Ergebnisse
64
Abb. 29: Signal des CCR4-Primers in qRT-PCR bei CASP- und Kontrollgruppe
In der CASP-Gruppe steigt die Fluoreszenz des Reportersignals (Delta RN) erst zu einem
späteren Zyklus an als in der unbehandelten Kontrollgruppe (WT). Das Signal des GAPDH-
Primers bleibt für beide Gruppen konstant. Aufgetragen ist die mittlere Fluoreszenz ∆ RN
gegen den Zyklus. Gezeigt sind die Ergebnisse isolierter RNA aus PBMCs gewonnen 20 h
nach CASP im Vergleich zu PBMCs unbehandelter Kontrolltiere.
Schon bei Betrachten der bloßen ∆RN-Werte (Abb. 29) liegt augenscheinlich
weniger CCR4-mRNA in den PBMCs der CASP-Tiere vor, als in den
gesunden Tieren. Nach zusätzlicher Einbeziehung der endogenen Kontrolle
GAPDH und Auswertung wird der Abfall von CCR4 noch deutlicher (p<0.001)
(Abb. 30, n = 9-10 pro Gruppe).
WT CASP0.0
0.5
1.0
1.5***
ct
Abb. 30: Abnahme der CCR4-Expression in qRT-PCR nach CASP
Page 73
Kapitel 4: Ergebnisse
65
20 h nach CASP kommt es zu einem signifikanten Abfall der CCR4-Expression auf mRNA-
Ebene (p<0.001). Untersucht wurde die RNA von PBMCs aus unbehandelten und septischen
(20 h CASP) C57BL/6-Mäusen. Die qRT-PCR wurde mit der ΔΔCt-Methode ausgewertet.
4.3.2 RT-qPCR muriner CD4+-Zellen
T-Helfer-Zellen (CD4+) wurden mit dem MACS-Kit aus der Milz isoliert. Die
Reinheit der CD4-Population betrug etwa 90%. Die RNA wurde mithilfe des
Quiagen-Mini Kit isoliert und anschließend mit dem NanoDrop untersucht. Für
die qRT-PCR wurde nur hochqualitative RNA verwendet, die keine
Verunreinigung mit Protein aufwiesen.
Vor Durchführung der qRT-PCR wurde die RNA in cDNA umgewandelt.
Verglichen wurde die cDNA und deren Gehalt an CCR4-mRNA von CASP-
Tieren (n = 14) mit der unbehandelten Kontrollgruppe (n = 12).
Die Fluoreszenzsignale des FAM-gekoppelten CCR4-Primers zeigen starke
Unterschiede zwischen den einzelnen Gruppen. In der CASP-Gruppe steigt
die Fluoreszenz erst zu einem viel späteren Zyklus (Abb. 31).
Abb. 31: Signal des CCR4-Primers in qRT-PCR bei CASP- und Kontrollgruppe
In der CASP-Gruppe steigt die Fluoreszenz des Reportersignals (Delta RN) erst zu einem
späteren Zyklus an als in der unbehandelten Kontrollgruppe. GAPDH wird als house-keeping
Gen in beiden Gruppen gleich exprimiert. Aufgetragen ist die mittlere Fluoreszenz ∆ RN gegen
den Zyklus.
Page 74
Kapitel 4: Ergebnisse
66
Nach Auswertung der PCR zeigt sich ein deutlicher Abfall der CCR4-mRNA in
den CASP-behandelten Tieren. T-Helfer-Zellen der septischen Tiere haben
eine signifikant geringere Expression von CCR4-mRNA im Zellkern (p<0.001)
(Abb. 32)
WT CASP0.0
0.5
1.0
1.5
2.0 ***
ct
Abb. 32: Abnahme der CCR4-Expression CD4+-Milzzellen in qRT-PCR nach CASP
20 h nach CASP kommt es zu einem signifikanten Abfall der CCR4-Expression auf mRNA-
Ebene (p<0.001). Untersucht wurde die RNA von CD4+-Milzzellen aus unbehandelten und
septischen C57BL/6-Mäusen. Die qRT-PCR wurde mit der ΔΔCt-Methode ausgewertet.
Sowohl PBMCs als auch CD4+-Zellen zeigen als Reaktion auf die CASP eine
verringerte Expression von CCR4 auf RNA-Ebene. Im Gegensatz dazu steht
die Hochregulation des Chemokinrezeptors in Thrombozyten nach CASP-
Induktion.
Page 75
Kapitel 5: Diskussion
67
5 Diskussion
In der vorliegenden Arbeit sollten die unterschiedlichen Funktionen von
Thrombozyten in der polymikrobiellen Sepsis untersucht werden. Zu diesem
Zweck wurden Versuche sowohl mit humanen als auch murinen Zellen
durchgeführt. Als anerkanntes Modell für die murine polymikrobielle Sepsis
wurde die Colon Ascendens Stent Peritonitis (CASP) sowohl bei C57BL/6 als
auch bei CCR4-/--Tieren angewandt. Humane Zellen wurden in vitro stimuliert.
5.1 In vitro Versuche mit humanen Zellen
In den letzten Jahren wurde durch viele Veröffentlichungen bekannt, dass die
Funktion der Thrombozyten über ihre Rolle in der Hämostase hinausreicht.
Gezeigt wurde, dass sie zusätzlich vor allem im angeborenen Immunsystem
von Bedeutung sind. Auf ihrer Oberfläche exprimieren sie die Toll-like-
Rezeptoren TLR 2, TLR 4 und TLR 9 [16, 40] und sind damit in der Lage,
Lipopolysaccharid (LPS) und andere bakterielle Strukturen zu binden. Eine
weitere Brücke der kernlosen Zellen zum Immunsystem ist ihre Expression
von P-Selektin. Das als Aktivierungsmarker genutzte Oberflächenprotein
befindet sich in den α-Granula der Thrombozyten und den Weibel-Palade-
Körperchen der Endothelzellen [41, 42]. Funktionell bedeutend ist P-Selektin
für das Rollen der Leukozyten auf dem Endothel der Gefäße vor der
Diapedese und damit das Einwandern der Immunzellen zum
Entzündungsgeschehen [43, 44]. Aktivierte Thrombozyten binden außerdem
Leukozyten über den Rezeptor P-Selektin Glykoprotein Ligand-1 (PSGL-1)
und vermitteln darüber den Durchtritt durch Hochendothelvenolen (HEVs) in
Lymphknoten und durch Mikrothromben an kleinen Gefäßschäden. Patienten,
die an einer Sepsis leiden und überflutet werden mit Pathogenen, zeigen
auch einen Anstieg der P-Selektin-Expression auf Thrombozyten [45, 46]. Wie
der Anstieg des Aktivierungsmarkers im Entzündungsprozess vermittelt wird,
ist in der Literatur widersprüchlich. Darum wurde um zu untersuchen, wie es
zu dem Anstieg des Aktivierungsmarkers durch den Entzündungsprozess
kommt, plättchenreiches Plasma (PRP) mit LPS und Lipoteichonsäure (LTA)
stimuliert. Es konnte sowohl nach LPS- als auch nach LTA-Stimulation kein
signifikanter Anstieg von P-Selektin auf den Thrombozyten beobachtet werden
Page 76
Kapitel 5: Diskussion
68
(Daten nur gezeigt für LPS). Sogar der sehr empfindliche Aktivierungsmarker
Pac-1, der den konvertierten funktionellen Fibrinogenrezeptor GPIIb/IIIa
spezifisch bindet, zeigte keine erhöhte Expression nach Stimulation. Auch
nach Vorinkubation mit dem Thrombozytenaktivator TRAP-6 konnte durch
LPS-Zugabe nur ein leichter Anstieg beider Aktivierungsmarker beobachtet
werden. Nach Ward et. al. ist LPS weder in der Lage via TLR-4 einen Anstieg
der P-Selektin-Expression auf Thrombozyten auszulösen noch Aggregation
und zytosolischen Calcium-Anstieg hervorzurufen [47]. TLR-4 und TLR-2
werden hier als funktionslose Überbleibsel der Megakaryozyten bezeichnet.
Andere Autoren sahen durch LPS-Stimulation von PRP zwar auch keinen
Anstieg in P-Selektin, wiesen dem TLR4 auf Thrombozyten dennoch stärkere
Bedeutung zu, da TLR4-/--Mäuse im Gegensatz zu Wildtyp-Mäusen nach LPS-
Gabe keine Thrombozytopenie aufwiesen [40]. Erstaunlicherweise wurde in
einer Quelle angegeben, dass LPS sogar zu einer Verminderung der
Thrombozytenaggregation führt, also die Expression des Fibrinogenrezeptors
und damit die Aktivierung verringert [48]. Im Kontrast dazu zeigen andere
Autoren, dass Thrombozyten neben TLR4 auch MyD88, MD2 und CD14 als
weitere beteiligte Proteine in der zugehörigen Signalkaskade exprimieren. Hier
wurde sowohl eine P-Selektin-Expression, als auch eine zusätzliche Erhöhung
des Aktivierungsmarkers nach Vorinkubation mit Thrombozytenstimulatoren
durch LPS-Gabe gesehen [49, 50]. Der Aktivierungs-Effekt durch LPS wurde
durch einen blockierenden TLR-4-Antikörper unterbunden, sodass ein
Signalweg über andere thrombozytäre Rezeptoren ausgeschlossen wurde.
Auch ein Interaktionseffekt mit anderen Zellen im Blut wurde durch die
Stimulation isolierter, gewaschener Thrombozyten unterbunden.
Die Widersprüchlichkeit der Literatur zeigt, wie empfindlich die Reaktion der
Thrombozyten auf äußere Faktoren ist. Es war zu überlegen, ob eine
Wiederholung mit größeren Gruppenzahlen die Aktivierung der Thrombozyten
im PRP durch LPS-Stimulation besser darstellen kann. Die durchgeführten
Experimente ließen jedoch darauf schließen, dass der Effekt sehr klein sein
würde und damit keine weiteren Versuche rechtfertigt. Außerdem wäre die
Relevanz in Anbetracht der diskutierten kontroversen Aussagen fraglich. Die
Bedeutung von TLR4 auf Thrombozyten wird vor allem im Zusammenspiel mit
anderen Zellen wichtig, wie es die nun zu diskutierenden Vollblutergebnisse
Page 77
Kapitel 5: Diskussion
69
zeigen werden. Hier führt LPS-Stimulation indirekt über andere Blutzellen zur
Aktivierung von Thrombozyten und zur Ausschüttung von Chemokinen wie
RANTES, PF4 und MIP-1α und damit zusätzlich zu einer Botschaft für andere
Immunzellen [19, 51, 52].
Im Gegensatz zum PRP, zeigte die Stimulation von Vollblut mit LPS einen
starken Anstieg von Thrombozyt-Leukozyt-Aggregaten und eine erhöhte P-
Selektin Expression der Thrombozyten. Beides sind Zeichen einer Aktivierung
und Degranulation der Plättchen. Die vermehrte Formation von Aggregaten ist
bereits von vielen Krankheitsbildern bekannt, wie Herzinfarkt, Diabetes oder
durch vaskuläre chirurgische Eingriffe [53-55]. Von besonderer Bedeutung ist
die Interaktion von Thrombozyten mit Granulozyten und Monozyten bei
Entzündung und Sepsis, da Thrombozyten ähnlich dem Endothel eine
Vermittlerrolle zwischen Leukozyten und Entzündungsherd einnehmen [56].
Die gefundene Erhöhung der Aggregate zwischen Thrombozyten und
Monozyten sowie Granulozyten nach LPS Stimulation wurde auch in der
Literatur beschrieben [56]. Sowohl die stärkere Bindung der Thrombozyten an
die Monozyten im Vergleich zu den Granulozyten, als auch eine nochmalige
Steigerung der Aggregatbildung nach Vorstimulation mit einem
Thrombozytenaktivator wurde von Autoren beschrieben [56]. Vermittelt wird
die Bindung über verschiedene Rezeptoren. Zuerst soll hier das ebenfalls
untersuchte P-Selektin genannt werden. P-Selektin aktivierter Thrombozyten
bindet den konstitutiv exprimierten PSGL-1-Rezeptor auf Granulozyten und
Monozyten [57]. Im Vergleich zum PRP war im Vollblut nach LPS-
Administration ein signifikanter Anstieg der P-Selektin-Expression zu sehen,
die Thrombozyten in den Aggregaten wurden also aktiviert. Dies lässt darauf
schließen, dass die Aktivierung der Plättchen nicht durch LPS direkt, sondern
durch einen indirekten Effekt mithilfe anderer Blutzellen vermittelt wurde. Eine
mögliche Hypothese wäre, dass die Bindung von LPS an Monozyten zur
Ausschüttung von Tissue Factor führt (TF) [58, 59] und TF die
Gerinnungskaskade auslöst, die in der Bildung von Thrombin endet. Thrombin
ist einer der stärksten Thrombozytenaktivatoren und aktiviert weitere
Thrombozyten im Vollblut über den protease-activated receptor (PAR) auf
ihrer Oberfläche [60].
Page 78
Kapitel 5: Diskussion
70
Da zwischen dem Anstieg der Aggregate (CD41-Expression) und der P-
Selektin-Expression eine relativ hohe Korrelation zu erkennen war (r= 0,817,
Daten nicht gezeigt), ist P-Selektin ein wesentlicher Rezeptor der
Thrombozyten, der die Interaktion mit PSGL-positiven Zellen vermittelt.
Allerdings lassen die Abweichungen und weniger korrelierenden Daten
vermuten, dass auch andere Rezeptoren bei der Vermittlung von Leukozyten
und Thrombozyten von Bedeutung sind. Diese Annahme lässt sich bestätigen
durch in der Literatur beschriebene Bindung zwischen Mac-1 auf Monozyten
und Granulozyten mit GPIbα, Bindungsstelle für den von Willebrand Faktor
(VWF) auf Thrombozyten [61, 62]. Im Gegensatz zur mäßigen Interaktion
zwischen P-Selektin und PSGL-1 ist die Bindung zwischen dem β2-Integrin
Mac-1 und GPIbα sehr stark und die beteiligten Rezeptoren werden erst nach
vorheriger Aktivierung der Leukozyten exprimiert [44, 61]. Die Aktivierung der
Immunzellen und damit die Expression von β2-Integrinen wird durch die oben
beschriebene Chemokin- und Zytokinausschüttung der Thrombozyten nach
LPS Stimulation hervorgerufen [51, 63]. Da Mac-1 zusätzlich Fibrinogen
bindet, kann darüber neben der direkten Bindung an Thrombozyten auch eine
indirekte Bindung zwischen Mac-1 und dem Fibrinogenrezeptor GpIIbIIIa
vermittelt werden [56]. Die LPS-Aktivierung löst also im Vollblut eine komplexe
Kaskade an Zell-Zell-Interaktionen aus, die von Diacovo et al. in folgendem
übersichtlichen Schema zusammengefasst wurden. Thrombozyten binden
Neutrophile in einem mehrstufigen Prozess und ermöglichen ähnlich dem
Endothel dadurch eine Passage zum Entzündungsherd. Im ersten Schritt des
Schemas kommt es zu einer schwachen, durch thrombozytäres P-Selektin
vermittelten Bindung und dadurch zum Rollen der Neutrophilen auf den
Plättchen. Im zweiten Schritt werden die Neutrophilen über Zytokine,
Chemokine oder Rezeptoren der Thrombozyten aktiviert. Dies führt im dritten
Schritt zu einer festen Bindung zwischen den beiden Zellen über das
induzierte Mac-1. Chemokin-vermittelt kommt es dann zuletzt zur Diapedese
durch die Thrombozytenschicht an den Entzündungsherd [44].
5.2 In vivo Versuche nach CASP
Nicht nur die Aktivierung von Thrombozyten, sondern auch die Abnahme der
Blutplättchenzahlen ist ein bekanntes pathologisches Phänomen bei
Page 79
Kapitel 5: Diskussion
71
septischen Patienten. 35-59% der erkrankten Patienten entwickeln eine
Thrombozytopenie [64, 65]. Auch im Tiermodell konnte nach CASP ein
signifikanter Abfall der Blutplättchen beobachtet werden, wie er auch schon für
die CLP-induzierte Sepsis beschrieben wurde [66]. Mögliche Ursachen für das
Sinken der Zellzahl können eine verringerte Produktion im Knochenmark, eine
erhöhte Destruktion und vermehrte Sequestration in kleinen Gefäßen sein
[67]. Die Immunhistochemie zur Ortung der Thrombozyten in den
verschiedenen Organen ergab vor allem die Ansammlung der Plättchen in der
Leber 20 Stunden nach CASP. Keine signifikanten Erhöhungen der
Thrombozyten wurden in Lunge, Milz und Niere (Daten für Niere nicht gezeigt)
beobachtet. Angaben in der Literatur, die ebenfalls Plättchenakkumulationen
in der Leber nachweisen konnten, stimmen mit diesen Ergebnissen überein
[68, 69]. Neben der Zunahme der Thrombozyten in der Leber, konnte auch
eine Erhöhung der Neutrophilen immunhistochemisch nachgewiesen werden.
Interessanterweise überlagerten sich beide Zellen optisch, waren also in
unmittelbarer Nähe zueinander. Dafür spricht, dass Leukozyten, vor allem
Neutrophile, nach CLP in die Leber einwandern und aktiviert werden [70, 71].
Singer et. al beschrieb die Adhäsion von Thrombozyten an das
Kapillarendothel der Leber in Abhängigkeit von der vorherigen Neutrophilen-
Einwanderung [70]. Von anderen Autoren wurde beschrieben, dass es durch
die LPS-Aktivierung von Thrombozyten via TLR-4 zur Bindung an die
Neutrophilen und zu deren Aktivierung kommt. Interaktionen der beiden
Zelltypen lösen die Bildung von Neutrophil Extracellular Traps (NETs), Netzen
aus fibrillärer Matrix und Chromatin, die das Einfangen von Bakterien
erleichtern, aus [52, 72]. Für diesen indirekten Weg über die Thrombozyten
spricht, dass Neutrophile allein nicht durch LPS zur NET-Bildung angeregt
werden konnten. Als Folge dieser Anreicherung von Thrombozyten und
Granulozyten in der Mikrozirkulation der Leber kommt es zu einer
verminderten Durchblutung des Organs und damit zur Nekrose [52, 71]. Dies
könnte eine Ursache für das Leberversagen während einer Sepsis sein.
Die leberspezifische Ansammlung von Thrombozyten nach CASP könnte
durch einen Rezeptor, der auf Hepatozyten exprimiert wird, verursacht
werden. Thrombozyten werden nach bakterieller Infektion durch die bakterielle
Neuraminidase in ihrer Oberflächenstruktur verändert. Deshalb können sie
Page 80
Kapitel 5: Diskussion
72
durch den sogenannten Ashwell-Rezeptor auf Hepatozyten erkannt werden,
reichern sich spezifisch in der Leber an und verschwinden aus der Zirkulation
Diese Beobachtungen können zusätzlich als Erklärung für die
Thrombozytopenie herangezogen werden [73].
Zwischen der CCR4-/-- und der C57BL/6-Maus wurde kein Unterschied in der
organspezifischen Thrombozytenanreicherung nach CASP gesehen. Auch die
Tiere, denen der CCR4-Rezeptor fehlte, zeigten 20 Stunden nach CASP eine
Thrombozytenanreicherung in der Leber und keine Veränderung der
Plättchenzahlen in Lunge und Milz im Vergleich zu unbehandelten knock-out-
Tieren. Der Chemokinrezeptor ist also vermutlich nicht an der Verteilung der
Thrombozyten in der Sepsis beteiligt, auch wenn Thrombozyten diesen
exprimieren [74]. Der in dieser Hinsicht fehlende Einfluss von CCR4 lässt sich
darauf zurückführen, dass weder Neutrophile noch Thrombozyten die
Liganden von CCR4, MDC und TARC, ausschütten oder produzieren.
Trotzdem beide Chemokine Thrombozytenaggregation und Calciumanstieg
auslösen, sind sie als Botenstoff eher für die Einwanderung von T-und B-
Zellen zum Entzündungsherd als für die Verteilung von Thrombozyten in der
Sepsis verantwortlich [31, 75, 76].
Im Vergleich zur Leber konnte in Milz und Lunge 20 h nach CASP keine
Akkumulation der Thrombozyten beobachtet werden. Im Gegensatz dazu
wurde von anderen Autoren allerdings nach LPS-Injektion eine Anhäufung der
Plättchen in der Lunge gefunden [40, 68, 69]. Als Erklärung für diesen
Widerspruch kann der Zeitpunkt der Organuntersuchung der Tiere
herangezogen werden. Während in dieser Arbeit alle Organe 20 Stunden nach
Sepsisinduktion untersucht wurden, wurde die Lunge in den anderen Quellen
nur wenige Minuten nach LPS-Injektion untersucht. Für zeitliche Abhängigkeit
spricht ein Schema von Ohtaki et. al., der die Verteilung der Thrombozyten
nach LPS-Gabe analysiert hat. Nur wenige Minuten nach LPS-Administration
häufen sich die Thrombozyten in der Lunge an und kehren anschließend in die
Zirkulation zurück. Nach etwa zehn Minuten akkumulieren sie in der Milz und
erst nach Stunden reichern sie sich in der Leber an. Diese Reihenfolge wurde
besonders nach i.v.-Injektion von hohen LPS-Dosen beobachtet. Injizierte man
geringe Dosen i.v. oder i.p., wurde nur die Einwanderung der Thrombozyten in
die Leber induziert [69, 77]. Diese Theorie bestätigt die beobachteten
Page 81
Kapitel 5: Diskussion
73
Ergebnisse der fehlenden Thrombozytenakkumulation in Lunge und Milz und
passt zu dem Anstieg in der Leber 20 Stunden nach CASP.
Zusätzlich zur Untersuchung nach 20 Stunden, wurde die Thrombozytenzahl
in der Leber auch 48 Stunden nach CASP überprüft. Hier zeigte sich immer
noch eine signifikante Erhöhung der Plättchen im Vergleich zu unbehandelten
Tieren, allerdings nahm ihre Anzahl im Vergleich zum früheren Zeitpunkt nach
20 Stunden in dem Organ deutlich ab.
Die Abnahme der Thrombozyten in der Leber nach 48 Stunden könnte drei
mögliche Ursachen haben. Zum einen könnte es zu einer erneuten
Umverteilung der Thrombozyten gekommen sein, wie es schon für die Lunge
beschrieben wurde. Die Thrombozyten gelangen also wieder in die Zirkulation
und wandern möglicherweise in ein anderes Organ. Als weitere Möglichkeit
könnte es nach 48 Stunden bereits zu einem Abbau der eingewanderten
Thrombozyten gekommen sein. Plausibler scheint jedoch die Annahme, dass
die Tiere, die 48 Stunden nach CASP überleben, schon 20 Stunden nach
Induktion der Sepsis eine geringere Thrombozytenanreicherung in der Leber
aufgewiesen haben. Die geringere Anzahl der Plättchen in der Mikrozirkulation
der Leber verhindert möglicherweise ein ischämisches Organversagen und
führt damit zum besseren Überleben dieser Tiere im Vergleich zu denen, die
20 Stunden nach CASP eine stärkere Thrombozytensequestration in der
Leber aufwiesen.
Als weitere Ursache der verminderten Thrombozytenzahl nach CASP wurde
die disseminierte intravasale Gerinnung (DIC) in Betracht gezogen. Dafür
wurde der Fibrinogengehalt im Plasma bestimmt. Wider erwarten kam es zu
einem signifikanten Anstieg des Fibrinogenplasmalevels 20 Stunden nach
CASP sowohl in der C57BL/6 als auch in der CCR4-/--Maus. Zur Ermittlung
einer DIC beim Patienten wird der sogenannte DIC-Score verwendet, erstellt
von der Internationalen Gesellschaft für Thrombose und Hämostase (ISTH).
Der Score beinhaltet die Thrombozytenzahl, den Fibrinogengehalt im Plasma,
Fibrinmarker (z.B. Fibrinspaltprodukte / D-Dimere, Fibrinmonomere) und die
Verlängerung der Prothrombinzeit in Sekunden (International normalized Ratio
= INR). Für die septischen Tiere wurde hier das Fibrinogen als Marker
ausgewählt und ein Absinken des Plasmagehalts durch die DIC erwartet.
Ursache für den unerwarteten Anstieg von Fibrinogen ist vor allem die
Page 82
Kapitel 5: Diskussion
74
gleichzeitige Funktion von Fibrinogen als Akute-Phase-Protein. Durch das
Trauma und die Entzündung nach CASP steigt Fibrinogen als Akute-Phase-
Protein an und verfälscht dessen Funktion als Marker für DIC. Weiterhin
bleiben die Plasmawerte trotz vorhandener gesteigerter Gerinnung von
Fibrinogen sehr lange konstant und erst bei sehr starker intravasaler
Koagulation kommt es zu einem Abfall [78, 79]. Auch in einer klinischen Studie
mit 40 septischen Patienten fand man nur einen Fall mit einem
Fibrinogengehalt, der unter der von der ISTH angegebenen Grenze lag [80].
Der Anstieg von Fibrinogen im Plasma ist kein Widerspruch, sondern nur
Zeichen der Entzündung nach CASP und hier kein gut gewählter Marker für
DIC.
5.3 CCR4-Regulation nach CASP
Chemokine und deren Rezeptoren spielen eine wichtige Rolle im
Zusammenspiel des Immunsystems. Der Chemokinrezeptor CCR4 und seine
Liganden CCL17 (TARC) und CCL22 (MDC) sind vor allem im
Zusammenhang mit Autoimmunkrankheiten wie atopische Dermatitis
untersucht worden. Ursache für die einseitige Forschung ist seine
vornehmliche Expression auf TH2- Zellen. Trotzdem ist CCR4 auch in andere
pathologische Mechanismen involviert. Die Abwesenheit des CCR4-Rezeptors
bringt sowohl nach LPS-Gabe als auch nach CLP oder CASP ein
Überlebensvorteil für die Tiere im Vergleich zu Wildtyp-Mäusen [30, 32, 81,
82].
Die CD4+-Milzzellen und peripheren mononukleären Blutzellen (PBMCs), die
in dieser Arbeit untersucht wurden, zeigten eine drastische Abnahme der
Expression von CCR4 auf mRNA-Ebene 20 Stunden nach CASP im Vergleich
zur unbehandelten Kontrollgruppe. Dabei verhalten sie sich konträr zu
Blutplättchen, die überraschenderweise als einzige bekannte Zellen CCR4 in
der Sepsis hoch regulieren. Dies konnte in einem Parallelprojekt unserer
Arbeitsgruppe gezeigt werden.
In der Literatur wurden ähnliche Regulationsmechanismen wie bei den CD4+-
Zellen und PBMCs für Leber, Milz, Thymus und mesenteriale Lymphknoten,
die 20 Stunden nach CASP untersucht wurden, beschrieben. Im Gegensatz
dazu war die Expression von CCR4 drei Stunden nach CASP in den
Page 83
Kapitel 5: Diskussion
75
jeweiligen Organen noch identisch zu den Werten der Kontrolltiere [30].
Interessanterweise zeigen PBMCs während der akuten Phase einer
allergischen Reaktion auf ein Medikament genau die entgegengesetzte
Regulation. CCR4 wird hier hoch reguliert [83]. Die Expression der beiden
Hauptliganden von CCR4, MDC und TARC, ist sowohl bei allergischen
Exanthemen der Haut als auch in septischer Milz 20 Stunden nach CASP auf
mRNA-Ebene erhöht [30, 83]. Die Regulation der beiden Chemokine ist
unabhängig von CCR4 und dessen Regulation, da die erhöhte Expression
auch in CCR4-/--Tieren nach CASP beobachtet wird [30].
Die Hochregulation der Chemokine CCL17 (TARC) und CCL22 (MDC) ist
einfach über die Bindung der bakteriellen Pathogen-assoziierten molekularen
Muster (PAMPs) an die pattern recognition receptors (PRR) zu erklären. LPS
als klassisches PAMP bindet beispielsweise an TLR4 und induziert damit die
Aktivierung des Transkriptionsfaktors NF-κB und die daraus resultierende
gesteigerte Transkription von Chemokinen und Zytokinen [32]. Die Regulation
von CCR4 lässt sich ungleich schwieriger erklären. Eine mögliche Hypothese
wäre die Hypoinflammation (CARS) als gegenregulatorische Maßnahme des
Körpers auf die systemische Hyperinflammation (SIRS). Beide Phasen kann
man während des Verlaufs einer Sepsis beobachten [84]. Die Ausschüttung
proinflammatorischer Zytokine, wie TNF bewirkt die reflektorische Sekretion
antiinflammatorische Zytokine, wie Il-10. Über negative Rückkopplung wird
außerdem eine weitere TNF-Sekretion verhindert. In einem analogen
Mechanismus könnte zusätzlich die Expression des Chemokinrezeptors CCR4
auf mRNA-Ebene verringert werden, so dass die Immunparalyse verstärkt
wird.
Zusammenfassend lässt sich sagen, dass Thrombozyten bei der Sepsis eine
wesentliche Rolle spielen. Thrombozytopenie, disseminierte intravasale
Gerinnung und Thrombozytenaktivierung erschweren das Krankheitsbild
septischer Patienten und könnten Angriffspunkte therapeutischer
Interventionen sein. Trotzdem Thrombozyten kernlos und mit einem
Durchmesser von 1,5-3 µm sehr klein sind, nehmen sie über Interaktion und
Kommunikation mit anderen Immunzellen maßgeblich Einfluss auf das
Krankheitsgeschehen. Sowohl in vitro, als auch in vivo konnte die Beteiligung
von Thrombozyten im inflammatorischen Geschehen gezeigt werden. In vitro
Page 84
Kapitel 5: Diskussion
76
interagierten Blutplättchen nach LPS-Stimulation mit Leukozyten, was die
Expression von Oberflächenmolekülen mit immunologischer Funktion
nachweist. Mithilfe des CASP-Modells wurde die Rolle der Thrombozyten in
der polymikrobiellen Sepsis am Mausmodell gezeigt. Die Einwanderung der
Thrombozyten vor allem in die Leber könnte hier eine mögliche Ursache für
ein Organversagen darstellen, da die Thromben und Thrombozyten-
Granulozyten-Aggregate die Mikrozirkulation beeinflussen und zur Nekrose
führen. Da das Organversagen einhergeht mit einer gesteigerten Letalität,
wäre die Verhinderung der Thrombozyteneinwanderung ein möglicher
therapeutischer Angriffspunkt.
Warum Thrombozyten viele Eigenschaften von Immunzellen aufweisen, lässt
sich anhand der Evolution erklären. Säugetiere sind die einzigen Lebewesen,
die polyploide Megakaryozyten mit kernlosen diskoiden Abkömmlingen als
Blutzellen aufweisen. Andere Wirbeltiere wie Vögel oder Fische besitzen
kernhaltige Thrombozyten, die als deren Hauptvertreter der weißen Blutzellen,
als Leukozyten fungieren. In wirbellosen Tieren findet man sogar nur eine
Blutzelle, den Amöbozyten, der trotz der 400 Millionen Jahre Evolution noch
erstaunliche Ähnlichkeit mit unseren Thrombozyten zeigt. Beide haben
beispielsweise eine diskoide Form, enthalten Granula und entwickeln
Pseudopodien nach Aktivierung [19, 85]. Diese Entwicklung erklärt die heute
noch vorhandene Funktion des Thrombozyts als Immunzelle und sollte
aufgrund der Menge an Plättchen sicherlich nicht unterschätzt werden.
Plättchen exprimieren und sezernieren eine große Anzahl an Zytokinen,
Mediatoren und Rezeptoren. Der Chemokinrezeptor CCR4 scheint dabei für
die Thrombozyten in der Sepsis von untergeordneter Bedeutung zu sein. Der
Überlebensvorteil, den CCR4-/--Mäuse in der Sepsis zeigen, ist wohl eher
durch das Fehlen des Rezeptors auf Leukozyten als auf Thrombozyten
bedingt.
Auch wenn immer neue Studien die vielseitige Funktion von Thrombozyten,
sowie deren Anpassungsfähigkeit aufzeigen, werden sie in ihrer Bedeutung
häufig noch unterschätzt. Daher sind dringend weitere Studien erforderlich um
die Komplexität dieser kleinen, kernlosen Zellen zu lernen und zu verstehen.
Page 85
Kapitel 6: Zusammenfassung
77
6 Zusammenfassung
In der vorliegenden Arbeit wurde in vitro und in vivo die Rolle der
Thrombozyten in der Sepsis untersucht. In vitro wurde sowohl humanes
plättchenreiches Plasma (PRP) als auch Vollblut mit LPS und TRAP-6, einem
Thrombinanalogon, stimuliert und eine mögliche Aktivierung der
Thrombozyten durchflusszytometrisch überprüft. Im PRP kam es zu einem
leichten Anstieg von P-Selektin, einem Aktivierungsmarker und
Adhäsionsmolekül auf Thrombozyten, nach Kostimulation von TRAP-6 und
LPS im Vergleich zur TRAP-6 behandelten Gruppe. Im Vollblut zeigte sich
nach LPS-Zugabe dagegen ein signifikanter Anstieg von Leukozyt-
Thrombozyt-Aggregaten und eine Zunahme der P-Selektin-Expression auf
den aggregatbildenden Thrombozyten.
In vivo wurde als etabliertes Sepsismodell die Colon Ascendens Stent
Peritonitis (CASP) durchgeführt. Weibliche C57BL/6-Mäuse wurden einer 16
G CASP unterzogen und nach 20 h untersucht. Unbehandelte Tiere dienten
als Kontrollgruppe. Es wurde ein signifikanter Abfall der Thrombozytenzahl im
peripheren Blut septischer Tiere beobachtet. Um die Ursache für diese
verminderte Plättchenanzahl zu finden, wurden zuerst immunhistochemische
Färbungen von Leber, Lunge, Niere und Milz angefertigt um dort mögliche
Thrombozytenanreicherungen zu entdecken. In diesen zeigte sich allerdings
nur in der Leber eine deutliche Zunahme von Thrombozyten nach CASP. In
Lunge, Niere und Milz war kein signifikanter Anstieg zu verzeichnen.
Als zweiter möglicher Grund für die Thrombozytopenie wurde die
disseminierte intravasale Gerinnung (DIC) als bekanntes Phänomen der
Sepsis in Betracht gezogen. Zur Überprüfung wurde der Fibrinogengehalt im
Plasma per ELISA bestimmt, da bei starker Gerinnung dessen Gehalt sinkt.
Dabei zeigte sich allerdings eine Zunahme von Fibrinogen im Plasma bei den
septischen Tieren im Vergleich zur unbehandelten Kontrollgruppe. Fibrinogen
sank also nicht im Sinne einer Verbrauchskoagulopathie, wie es zu erwarten
wäre, sondern stieg im Sinne eines Akute-Phase-Proteins.
Weiterhin wurde der Einfluss des Chemokinrezeptors CCR4 in der Sepsis
untersucht, da CCR4-/--Mäuse im Vergleich zu C57BL/6-Tieren nach CASP
einen Überlebensvorteil haben. Zuerst wurde die Expression von CCR4 auf
Page 86
Kapitel 6: Zusammenfassung
78
mRNA-Ebene in mononukleären Zellen des peripheren Blutes und CD4+-
Zellen der Milz 20 h nach CASP gemessen. In beiden Zellpopulationen zeigte
sich im Vergleich zur unbehandelten Kontrollgruppe eine signifikant
verringerte Transkription von CCR4. Da auch Thrombozyten CCR4
exprimieren, wurde sowohl die Immunhistochemie, als auch der ELISA bei
CCR4-/--Tieren nach CASP durchgeführt um mögliche Unterschiede zur
Wildtyp-Maus festzustellen. In beidem war allerdings kein Unterschied
zwischen C57BL/6- und CCR4-/--Tieren zu sehen.
Da Thrombozyten in vitro durch LPS aktiviert wurden, vor allem im
Zusammenspiel mit anderen Blutzellen und septische Tiere sowohl eine
Thrombozytopenie als auch eine Plättchenakkumulation in der Leber zeigten,
sind die Plättchen im Krankheitsverlauf der Sepsis von Bedeutung. Der
Chemokinrezeptor CCR4 scheint im Zusammenhang mit Thrombozyten in der
Sepsis von untergeordneter Bedeutung zu sein. In anderen Immunzellen, vor
allem T-Helfer-Zellen, wird dessen Expression im Zellkern im Verlauf der
Sepsis reguliert und könnte damit Einfluss auf den Krankheitsverlauf haben.
Page 87
79
Literaturverzeichnis
1. Martin, G.S., et al., The epidemiology of sepsis in the United States from 1979
through 2000. N Engl J Med, 2003. 348(16): p. 1546-54.
2. Angus, D.C., et al., Epidemiology of severe sepsis in the United States:
analysis of incidence, outcome, and associated costs of care. Crit Care Med,
2001. 29(7): p. 1303-10.
3. Engel, C., et al., Epidemiology of sepsis in Germany: results from a national
prospective multicenter study. Intensive Care Med, 2007. 33(4): p. 606-18.
4. Moerer, O. and M. Quintel, [Sepsis in adult patients - definitions, epidemiology
and economic aspects]. Internist (Berl), 2009. 50(7): p. 788, 790-4, 796-8.
5. Rivers, E., et al., Early goal-directed therapy in the treatment of severe sepsis
and septic shock. N Engl J Med, 2001. 345(19): p. 1368-77.
6. Hotchkiss, R.S. and I.E. Karl, The pathophysiology and treatment of sepsis. N
Engl J Med, 2003. 348(2): p. 138-50.
7. Hauber, H.P. and P. Zabel, [Pathophysiology and pathogens of sepsis].
Internist (Berl), 2009. 50(7): p. 779-80, 782-4, 786-7.
8. Okazaki, Y. and A. Matsukawa, Pathophysiology of sepsis and recent patents
on the diagnosis, treatment and prophylaxis for sepsis. Recent Pat Inflamm
Allergy Drug Discov, 2009. 3(1): p. 26-32.
9. Denis, M.M., et al., Escaping the nuclear confines: signal-dependent pre-
mRNA splicing in anucleate platelets. Cell, 2005. 122(3): p. 379-91.
10. Schwertz, H., et al., Anucleate platelets generate progeny. Blood, 2010.
115(18): p. 3801-9.
11. Schwertz, H., et al., Signal-dependent splicing of tissue factor pre-mRNA
modulates the thrombogenicity of human platelets. J Exp Med, 2006. 203(11):
p. 2433-40.
12. Weyrich, A.S., et al., Protein synthesis by platelets: historical and new
perspectives. J Thromb Haemost, 2009. 7(2): p. 241-6.
13. Rendu, F. and B. Brohard-Bohn, The platelet release reaction: granules'
constituents, secretion and functions. Platelets, 2001. 12(5): p. 261-73.
14. Rodak, B.F., Hematology: Clinical Principles and Applications. 2007: W.B.
Saunders Company.
Page 88
80
15. Michelson, A.D., Platelets. 2006: Oxford Elsevier Ltd.
16. Aslam, R., et al., Platelet Toll-like receptor expression modulates
lipopolysaccharide-induced thrombocytopenia and tumor necrosis factor-alpha
production in vivo. Blood, 2006. 107(2): p. 637-41.
17. Beaulieu, L.M. and J.E. Freedman, The role of inflammation in regulating
platelet production and function: Toll-like receptors in platelets and
megakaryocytes. Thromb Res, 2010. 125(3): p. 205-9.
18. Glusa, E., et al., Thrombin receptor activating peptide-induced cellular effects:
comparative studies on human platelet activation and endothelium-dependent
relaxation of porcine pulmonary arteries. Agents Actions Suppl, 1995. 45: p.
303-7.
19. Kehrel, B.E. and K. Jurk, Platelets at the interface between hemostasis and
innate immunity. Transfusion Medicine and Hemotherapy, 2004. 31(6): p. 379-
386.
20. Jennings, L.K., Mechanisms of platelet activation: need for new strategies to
protect against platelet-mediated atherothrombosis. Thromb Haemost, 2009.
102(2): p. 248-57.
21. Frojmovic, M.M. and J.G. Milton, Human platelet size, shape, and related
functions in health and disease. Physiol Rev, 1982. 62(1): p. 185-261.
22. Halvorsen, H., J.O. Olsen, and B. Osterud, Granulocytes enhance LPS-
induced tissue factor activity in monocytes via an interaction with platelets. J
Leukoc Biol, 1993. 54(4): p. 275-82.
23. Osterud, B., L.V. Rao, and J.O. Olsen, Induction of tissue factor expression in
whole blood: lack of evidence for the presence of tissue factor expression in
granulocytes. Thromb Haemost, 2000. 83(6): p. 861-7.
24. Taylor, F.B., Jr., et al., Towards definition, clinical and laboratory criteria, and a
scoring system for disseminated intravascular coagulation. Thromb Haemost,
2001. 86(5): p. 1327-30.
25. Levi, M., Platelets. Crit Care Med, 2005. 33(12 Suppl): p. S523-5.
26. Schouten, M., et al., Inflammation, endothelium, and coagulation in sepsis. J
Leukoc Biol, 2008. 83(3): p. 536-45.
27. Opal, S.M. and C.T. Esmon, Bench-to-bedside review: functional relationships
between coagulation and the innate immune response and their respective
roles in the pathogenesis of sepsis. Crit Care, 2003. 7(1): p. 23-38.
Page 89
81
28. Dempfle, C.E., Coagulopathy of sepsis. Thrombosis and Haemostasis, 2004.
91(2): p. 213-224.
29. Bone, R.C., et al., Definitions for sepsis and organ failure and guidelines for
the use of innovative therapies in sepsis. The ACCP/SCCM Consensus
Conference Committee. American College of Chest Physicians/Society of
Critical Care Medicine. Chest, 1992. 101(6): p. 1644-55.
30. Traeger, T., et al., Detrimental role of CC chemokine receptor 4 in murine
polymicrobial sepsis. Infect Immun, 2008. 76(11): p. 5285-93.
31. Abi-Younes, S., M. Si-Tahar, and A.D. Luster, The CC chemokines MDC and
TARC induce platelet activation via CCR4. Thromb Res, 2001. 101(4): p. 279-
89.
32. Ness, T.L., et al., CCR4 is a key modulator of innate immune responses. J
Immunol, 2006. 177(11): p. 7531-9.
33. Philipp M. Lepper, M.T., Medizinische Mikrobiologie und Infektiologie, ed. S.B.
Heidelberg. Vol. 6. 2009.
34. Maier, S., et al., [Special aspects of abdominal sepsis]. Chirurg, 2005. 76(9):
p. 829-36.
35. Wichterman, K.A., A.E. Baue, and I.H. Chaudry, Sepsis and septic shock--a
review of laboratory models and a proposal. J Surg Res, 1980. 29(2): p. 189-
201.
36. Zantl, N., et al., Essential role of gamma interferon in survival of colon
ascendens stent peritonitis, a novel murine model of abdominal sepsis. Infect
Immun, 1998. 66(5): p. 2300-9.
37. Bennett, J.S., et al., Interaction of fibrinogen with its platelet receptor.
Differential effects of alpha and gamma chain fibrinogen peptides on the
glycoprotein IIb-IIIa complex. J Biol Chem, 1988. 263(26): p. 12948-53.
38. Winokur, R. and J.H. Hartwig, Mechanism of shape change in chilled human
platelets. Blood, 1995. 85(7): p. 1796-804.
39. McEver, R.P., Properties of GMP-140, an inducible granule membrane protein
of platelets and endothelium. Blood Cells, 1990. 16(1): p. 73-80; discussion
80-3.
40. Andonegui, G., et al., Platelets express functional Toll-like receptor-4. Blood,
2005. 106(7): p. 2417-23.
Page 90
82
41. Stenberg, P.E., et al., A platelet alpha-granule membrane protein (GMP-140)
is expressed on the plasma membrane after activation. J Cell Biol, 1985.
101(3): p. 880-6.
42. McEver, R.P., et al., GMP-140, a platelet alpha-granule membrane protein, is
also synthesized by vascular endothelial cells and is localized in Weibel-
Palade bodies. J Clin Invest, 1989. 84(1): p. 92-9.
43. Diacovo, T.G., et al., Platelet-mediated lymphocyte delivery to high endothelial
venules. Science, 1996. 273(5272): p. 252-5.
44. Diacovo, T.G., et al., Neutrophil rolling, arrest, and transmigration across
activated, surface-adherent platelets via sequential action of P-selectin and
the beta 2-integrin CD11b/CD18. Blood, 1996. 88(1): p. 146-57.
45. Jacoby, R.C., et al., Platelet activation and function after trauma. J Trauma,
2001. 51(4): p. 639-47.
46. Ogura, H., et al., Activated platelets enhance microparticle formation and
platelet-leukocyte interaction in severe trauma and sepsis. J Trauma, 2001.
50(5): p. 801-9.
47. Ward, J.R., et al., Agonists of toll-like receptor (TLR)2 and TLR4 are unable to
modulate platelet activation by adenosine diphosphate and platelet activating
factor. Thromb Haemost, 2005. 94(4): p. 831-8.
48. Hashimoto, K., et al., Aggregation and microparticle production through toll-
like receptor 4 activation in platelets from recently menopausal women. J
Cardiovasc Pharmacol, 2009. 54(1): p. 57-62.
49. Zhang, G., et al., Lipopolysaccharide stimulates platelet secretion and
potentiates platelet aggregation via TLR4/MyD88 and the cGMP-dependent
protein kinase pathway. J Immunol, 2009. 182(12): p. 7997-8004.
50. Shashkin, P.N., et al., Lipopolysaccharide is a direct agonist for platelet RNA
splicing. J Immunol, 2008. 181(5): p. 3495-502.
51. Cognasse, F., et al., Toll-like receptor 4 ligand can differentially modulate the
release of cytokines by human platelets. Br J Haematol, 2008. 141(1): p. 84-
91.
52. Clark, S.R., et al., Platelet TLR4 activates neutrophil extracellular traps to
ensnare bacteria in septic blood. Nat Med, 2007. 13(4): p. 463-9.
Page 91
83
53. Tefferi, A. and M. Elliott, Thrombosis in myeloproliferative disorders:
prevalence, prognostic factors, and the role of leukocytes and JAK2V617F.
Semin Thromb Hemost, 2007. 33(4): p. 313-20.
54. Elalamy, I., et al., Circulating platelet-leukocyte aggregates: a marker of
microvascular injury in diabetic patients. Thromb Res, 2008. 121(6): p. 843-8.
55. McEver, R.P., P-selectin and PSGL-1: exploiting connections between
inflammation and venous thrombosis. Thromb Haemost, 2002. 87(3): p. 364-5.
56. Stahl, A.L., et al., Shiga toxin and lipopolysaccharide induce platelet-leukocyte
aggregates and tissue factor release, a thrombotic mechanism in hemolytic
uremic syndrome. PLoS One, 2009. 4(9): p. e6990.
57. Mehta, P., et al., Soluble monomeric P-selectin containing only the lectin and
epidermal growth factor domains binds to P-selectin glycoprotein ligand-1 on
leukocytes. Blood, 1997. 90(6): p. 2381-9.
58. Steiner, S., et al., Simvastatin blunts endotoxin-induced tissue factor in vivo.
Circulation, 2005. 111(14): p. 1841-6.
59. Kalsch, T., et al., Endotoxin-induced effects on platelets and monocytes in an
in vivo model of inflammation. Basic Res Cardiol, 2007. 102(5): p. 460-6.
60. Marjoram, R.J., et al., Suboptimal activation of protease-activated receptors
enhances alpha2beta1 integrin-mediated platelet adhesion to collagen. J Biol
Chem, 2009. 284(50): p. 34640-7.
61. Ehlers, R., et al., Targeting platelet-leukocyte interactions: identification of the
integrin Mac-1 binding site for the platelet counter receptor glycoprotein
Ibalpha. J Exp Med, 2003. 198(7): p. 1077-88.
62. Bergmeier, W., et al., The role of platelet adhesion receptor GPIbalpha far
exceeds that of its main ligand, von Willebrand factor, in arterial thrombosis.
Proc Natl Acad Sci U S A, 2006. 103(45): p. 16900-5.
63. McNicol, A. and S.J. Israels, Beyond hemostasis: the role of platelets in
inflammation, malignancy and infection. Cardiovasc Hematol Disord Drug
Targets, 2008. 8(2): p. 99-117.
64. Yan, S.B., et al., Low levels of protein C are associated with poor outcome in
severe sepsis. Chest, 2001. 120(3): p. 915-22.
65. Brun-Buisson, C., et al., Incidence, risk factors, and outcome of severe sepsis
and septic shock in adults. A multicenter prospective study in intensive care
units. French ICU Group for Severe Sepsis. JAMA, 1995. 274(12): p. 968-74.
Page 92
84
66. Lerolle, N., et al., von Willebrand factor is a major determinant of ADAMTS-13
decrease during mouse sepsis induced by cecum ligation and puncture. J
Thromb Haemost, 2009. 7(5): p. 843-50.
67. Aird, W.C., The hematologic system as a marker of organ dysfunction in
sepsis. Mayo Clin Proc, 2003. 78(7): p. 869-81.
68. Shibazaki, M., et al., Complement-dependent accumulation and degradation of
platelets in the lung and liver induced by injection of lipopolysaccharides.
Infect Immun, 1999. 67(10): p. 5186-91.
69. Shibazaki, M., M. Nakamura, and Y. Endo, Biphasic, organ-specific, and
strain-specific accumulation of platelets induced in mice by a
lipopolysaccharide from Escherichia coli and its possible involvement in shock.
Infect Immun, 1996. 64(12): p. 5290-4.
70. Singer, G., et al., Platelet recruitment in the murine hepatic microvasculature
during experimental sepsis: role of neutrophils. Microcirculation, 2006. 13(2):
p. 89-97.
71. Gawaz, M., et al., Platelet activation and interaction with leucocytes in patients
with sepsis or multiple organ failure. Eur J Clin Invest, 1995. 25(11): p. 843-51.
72. Logters, T., et al., The clinical value of neutrophil extracellular traps. Med
Microbiol Immunol, 2009. 198(4): p. 211-9.
73. Grewal, P.K., et al., The Ashwell receptor mitigates the lethal coagulopathy of
sepsis. Nat Med, 2008. 14(6): p. 648-55.
74. Clemetson, K.J., et al., Functional expression of CCR1, CCR3, CCR4, and
CXCR4 chemokine receptors on human platelets. Blood, 2000. 96(13): p.
4046-54.
75. Imai, T., et al., The T cell-directed CC chemokine TARC is a highly specific
biological ligand for CC chemokine receptor 4. J Biol Chem, 1997. 272(23): p.
15036-42.
76. Imai, T., et al., Macrophage-derived chemokine is a functional ligand for the
CC chemokine receptor 4. J Biol Chem, 1998. 273(3): p. 1764-8.
77. Ohtaki, Y., et al., In vivo platelet response to lipopolysaccharide in mice:
proposed method for evaluating new antiplatelet drugs. Thromb Res, 2002.
108(5-6): p. 303-9.
78. Levi, M., Disseminated intravascular coagulation. Crit Care Med, 2007. 35(9):
p. 2191-5.
Page 93
85
79. Lissalde-Lavigne, G., et al., Simple coagulation tests improve survival
prediction in patients with septic shock. J Thromb Haemost, 2008. 6(4): p.
645-53.
80. Voves, C., W.A. Wuillemin, and S. Zeerleder, International Society on
Thrombosis and Haemostasis score for overt disseminated intravascular
coagulation predicts organ dysfunction and fatality in sepsis patients. Blood
Coagul Fibrinolysis, 2006. 17(6): p. 445-51.
81. Ishii, M., et al., CC chemokine receptor 4 modulates Toll-like receptor 9-
mediated innate immunity and signaling. Eur J Immunol, 2008. 38(8): p. 2290-
302.
82. Chvatchko, Y., et al., A key role for CC chemokine receptor 4 in
lipopolysaccharide-induced endotoxic shock. J Exp Med, 2000. 191(10): p.
1755-64.
83. Tapia, B., et al., Up-regulation of CCL17, CCL22 and CCR4 in drug-induced
maculopapular exanthema. Clin Exp Allergy, 2007. 37(5): p. 704-13.
84. Hoflich, C. and H.D. Volk, [Immunomodulation in sepsis]. Chirurg, 2002.
73(11): p. 1100-4.
85. Weyrich, A.S., S. Lindemann, and G.A. Zimmerman, The evolving role of
platelets in inflammation. J Thromb Haemost, 2003. 1(9): p. 1897-905.
Page 94
86
Verzeichnis der Abbildungen und Tabellen
Abbildungen
Abb. 1: Schematische Darstellung eines Thrombozyts mit all seinen ................
……….Organellen ............................................................................................ 3
Abb. 2: Änderung der Thrombozytenform nach Aktivierung ............................. 6
Abb. 3: Typische Thrombozytenpopulation in der Durchflusszytometrie ........ 39
Abb. 4: CD62P-Expression nach Stimulation von PRP mit unterschiedlichen
. TRAP-6 Konzentrationen ................................................................... 40
Abb. 5: Histogramme der CD62P- und PAC-1-Fluoreszenz nach Stimulation .
. mit TRAP-6 und LPS .......................................................................... 43
Abb. 6: Mittlere Fluoreszenz von PAC-1 und P-Selektin nach Stimulation ..... 44
Abb. 7: Durchflusszytometrische Darstellung der Zellen im Vollblut .............. 46
Abb. 8: Zunahme der Thrombozyten-Granulozyten-Aggregate ..................... 46
Abb. 9: Unterschiedliche Expression des Thrombozytenmarkers CD41 in der
. Granulozytenpopulation nach Stimulation mit LPS und TRAP-6 ........ 47
Abb. 10: CD62P -Expression der Thrombozyten in der Granulozytenpopulation
....................................................................................................................... 48
Abb. 11: Unterschiedliche Aktivierung der Thrombozyten in der .
. Granulozytenpopulation ................................................................... 48
Abb. 12: Zunahme der Thrombozyten-Monozyten-Aggregate nach LPS-.
…………Stimulation ....................................................................................... 49
Abb. 13: CD41-Expression in der Monozyten-Population nach LPS und
. TRAP-6 Zugabe ............................................................................... 50
Abb. 14: CD62P-Expression der Thrombozyten in der Monozytenpopulation 50
Abb. 15: Unterschiedliche Aktivierung der Thrombozyten in der .
…………Monozytenpopulation ....................................................................... 51
Abb. 16: Änderung der Thrombozytenzahl nach CASP ................................. 52
Abb. 17: Immunhistologische Färbungen von Lunge und Milz ....................... 54
Abb. 18: Änderung der Thrombozytenzahl in der Lunge ................................ 55
Abb. 19: Änderung der Thrombozytenzahl in der Milz ................................... 55
Abb. 20: Immunhistologische Färbungen der Leber ...................................... 56
Page 95
87
Abb. 21: Änderung der Thrombozytenzahl in der Leber ................................. 57
Abb. 22: Immunhistochemische Anfärbung von P-Selektin und CD42c auf
………... Leber ............................................................................................... 58
Abb. 23: Zunahme der P-Selektin-Expression in der Leber nach CASP ........ 58
Abb. 24: Immunhistochemische Anfärbung von CD42c und Gr-1 in der Leber
....................................................................................................................... 59
Abb. 25: Zunahme der Granulozyten in der Leber 20 h nach CASP .............. 60
Abb. 26: Standardreihe für den Fibrinogen-ELISA ......................................... 61
Abb. 27: Zunahme von Fibrinogen im Plasma 20h nach CASP ..................... 61
Abb. 28: Abnahme der Leukozyten 20 h nach CASP..................................... 63
Abb. 29: Signal des CCR4-Primers in qRT-PCR bei CASP- und Kontrollgruppe
....................................................................................................................... 64
Abb. 30: Abnahme der CCR4-Expression in qRT-PCR nach CASP .............. 64
Abb. 31: Signal des CCR4-Primers in qRT-PCR bei CASP- und Kontrollgruppe
....................................................................................................................... 65
Abb. 32: Abnahme der CCR4-Expression CD4+-Milzzellen in qRT-PCR nach
………...CASP ............................................................................................... 66
Tabellen
Tabelle 1: Versuchsaufbau der Stimulation von PRP..................................... 42
Tabelle 2: Versuchsaufbau der Stimulation von Vollblut ................................ 45
Page 96
Eidesstattliche Erklärung
Hiermit erkläre ich, dass ich die vorliegende Dissertation selbstständig verfasst
und keine anderen als die angegebenen Hilfsmittel verwendet habe.
Die Dissertation ist bisher keiner anderen Fakultät vorgelegt worden.
Ich erkläre, dass ich bisher kein Promotionsverfahren erfolglos beendet habe
und dass eine Aberkennung eines bereits erworbenen Doktorgrades nicht
vorliegt.
Datum Unterschrift
Page 97
Lebenslauf
Persönliche Daten
Schulbildung
08/93-07/99
08/99-06/06
Studium
10/06
04/09-04/10
04/09-04/11
09/09/09-13/09/09
08/03/10-13/03/10
Manja Appelt
Roald-Amundsen Str. 12
17493 Greifswald
[email protected]
Geb. am 08.12.1986 in Schwedt
Grundschule 6 Schwedt
Carl-Friedrich-Gauss-Gymnasium
Schwedt
Abiturnote: 1,2
Aufnahme des Studiums der
Humanmedizin an der Ernst-Moritz-
Arndt Universität Greifswald
Physikumsnote: 1,5
Laborjahr im Rahmen meiner
Doktorarbeit in der Chirurgie
Mitglied im Graduiertenkolleg 840
Projekt E1 „The effect of CCR4 on
immuno-competent cells in
inflammation”
Aktive Kongressteilnahme
Weimar Sepsis Update 2009
Aktive Kongressteilnahme
Page 98
ab 11/10
Berufstätigkeit
03/07-04/07
08/07-10/07
04/08-07/08
10/08-07/11
07/10-08/10
08/10-09/10
09/10-10/10
Auslandsaufenthalte
01/03-06/03
„Trauma, shock, inflammation and
sepsis – TSIS 2010“
Weiterführung des Medizinstudiums
Pflegepraktikum in der
Gastroenterologie Greifswald
Pflegepraktikum in der plastischen
Chirurgie
des Vivantes Klinikum Berlin
Anstellung an der EMAU als
Chemietutorin
Anstellung an der EMAU als
Biochemietutorin
Famulatur im Franziskus
Krankenhaus Berlin
-Innere Abteilung-
Famulatur im Asklepios Klinikum
Uckermark
-Abteilung für
Orthopädie/Unfallchirurgie-
Famulatur im Asklepios Klinikum
Uckermark
-Abteilung für Neurologie-
Highschoolyear in den USA, Missouri
Page 99
Danksagung
Diese Arbeit hätte ohne die Hilfe und tatkräftige Unterstützung von vielen
Seiten nicht verwirklicht werden können.
Zu allererst gilt ein besonderer Dank Herrn Prof. Dr. med. Claus-Dieter-
Heidecke und Prof. Dr. med. Stefan Maier, die mich in ihre Arbeitsgruppe
aufgenommen haben und die ausgiebige Arbeit an diesem interessanten
Thema ermöglicht haben. Weiterhin danken möchte ich auch den Mitarbeitern
der Arbeitsgruppe Frau Dr. med. Alexandra Busemann, Frau Dr. med. Pia
Körner, Frau Dr. med. Katharina Cziupka, Herrn Dr. med. Tobias Traeger und
Herrn Dr. med. Wolfram Keßler, die für Anregungen, Diskussionen und
Verbesserungsvorschläge in den Laborbesprechungen gesorgt haben. Ein
besonderer Dank gilt hier Stephan Diedrich, der sich im Labor viel Zeit für
Gespräche genommen hat und mich in die operativen Techniken eingeführt
hat.
Außerdem danke ich ganz besonders Frau Antje Janetzko und Frau Doreen
Biedenweg für ihre große Unterstützung und ihre Geduld in der Durchführung
vieler Experimente. Auch Frau Birgitt Fürll aus der Abteilung für
Transfusionsmedizin, die mir mit Ideen und technischem Wissen sehr
geholfen hat, möchte ich herzlich danken.
Danken möchte ich auch meinen lieben Mitdoktoranden Laura Klene, Michael
Nielson und Christian Klöcker für eine sehr angenehme Arbeitsatmosphäre
und die gegenseitige Unterstützung. Ohne sie wäre das Jahr im Labor sicher
um viele schöne Momente ärmer gewesen.
Der Deutschen Forschungsgemeinschaft gilt mein Dank für ein Stipendium
und wissenschaftliche Beihilfe im Rahmen des Graduiertenkollegs 840 Host-
pathogen interactions in generalized bacterial infections. Unter der Leitung von
Prof. Dr. med. Barbara Bröker wurden mir dort das wissenschaftliche Arbeiten,
der wissenschaftliche Austausch und die Teilnahme an Kongressen näher
gebracht und ermöglicht.
Ein riesiges Dankeschön möchte ich auch an meine Eltern aussprechen, die
mich in jeder Hinsicht im Studium und in meiner Doktorarbeit unterstützt
haben. Neben meinen Eltern gilt auch meinem Freund Torben ein besonderer
Page 100
Dank für seine geduldige, liebevolle Unterstützung und den ermutigenden
Beistand in jeder Situation.














































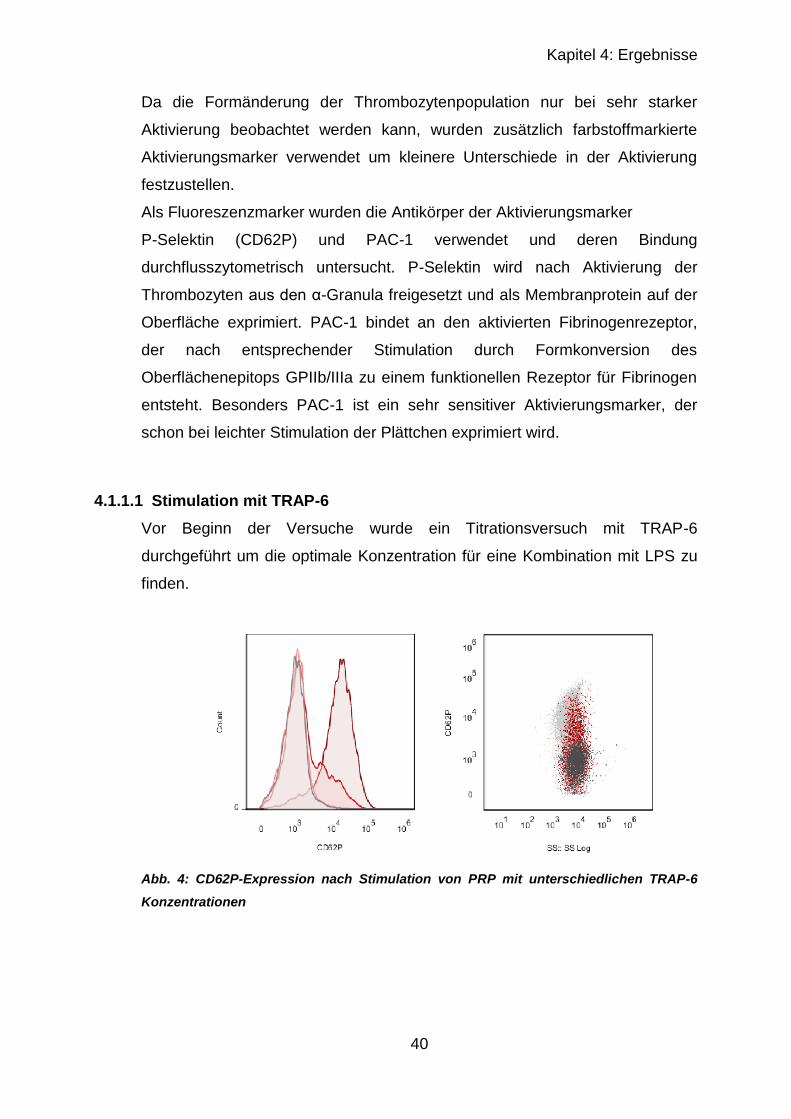

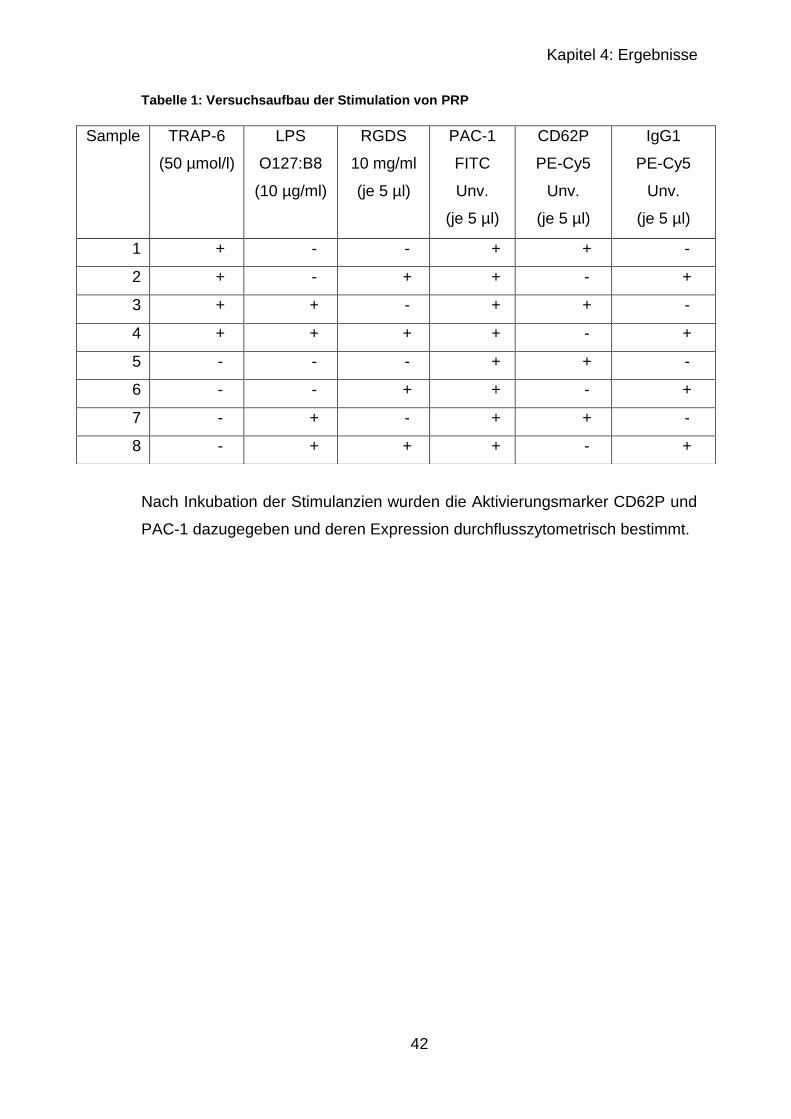
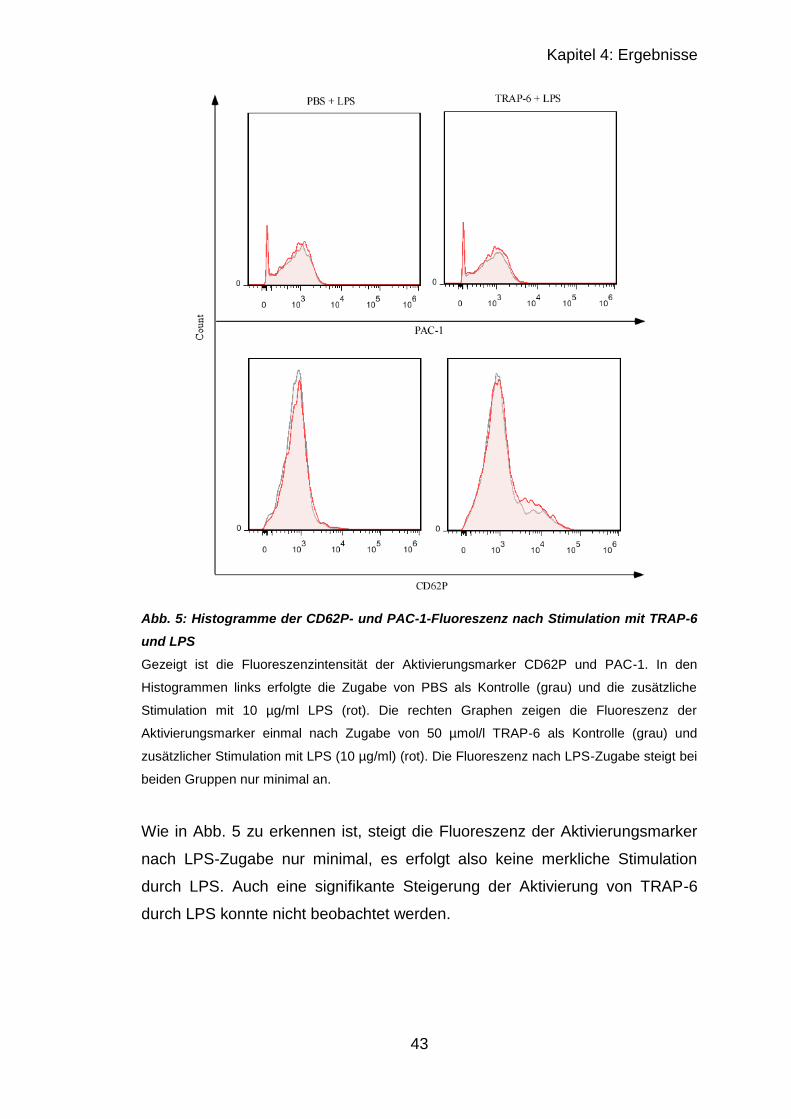
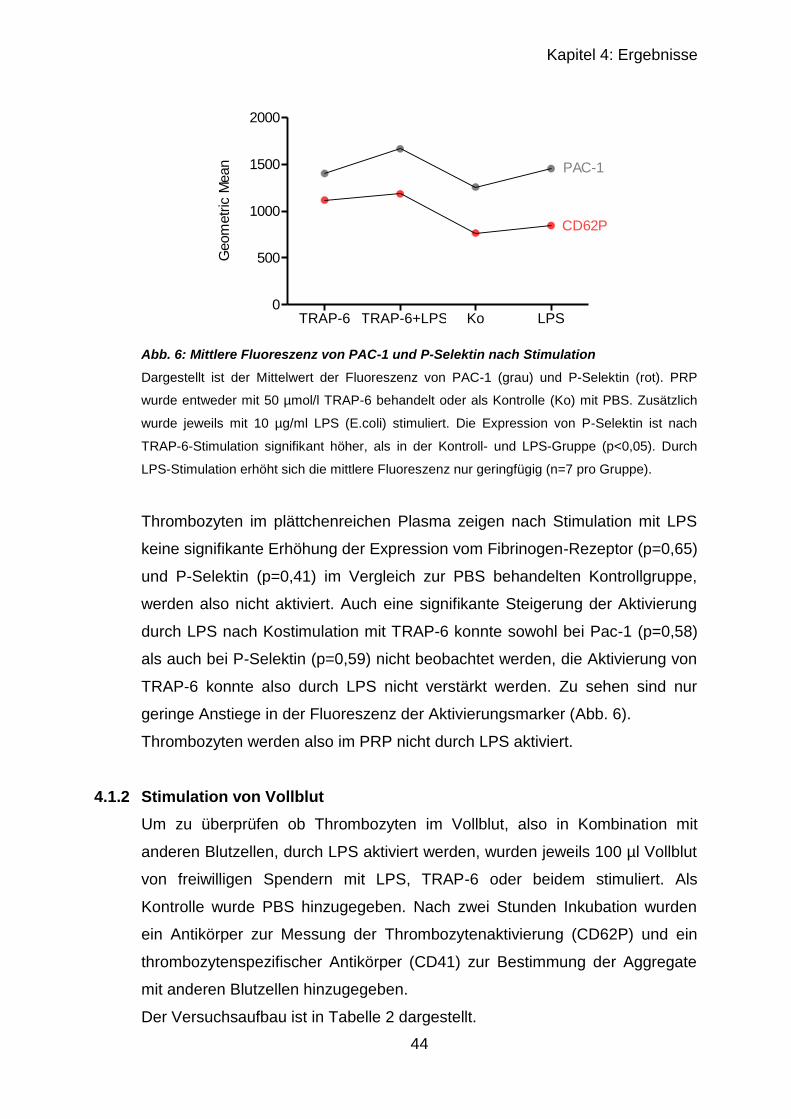
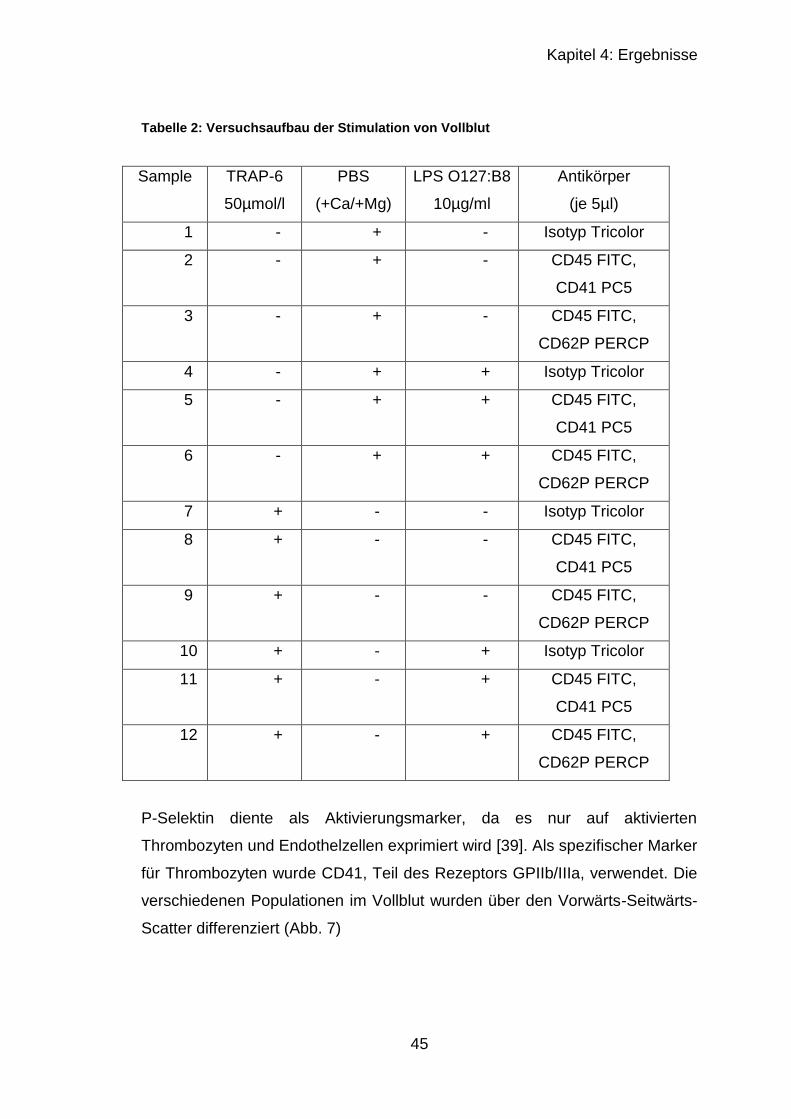
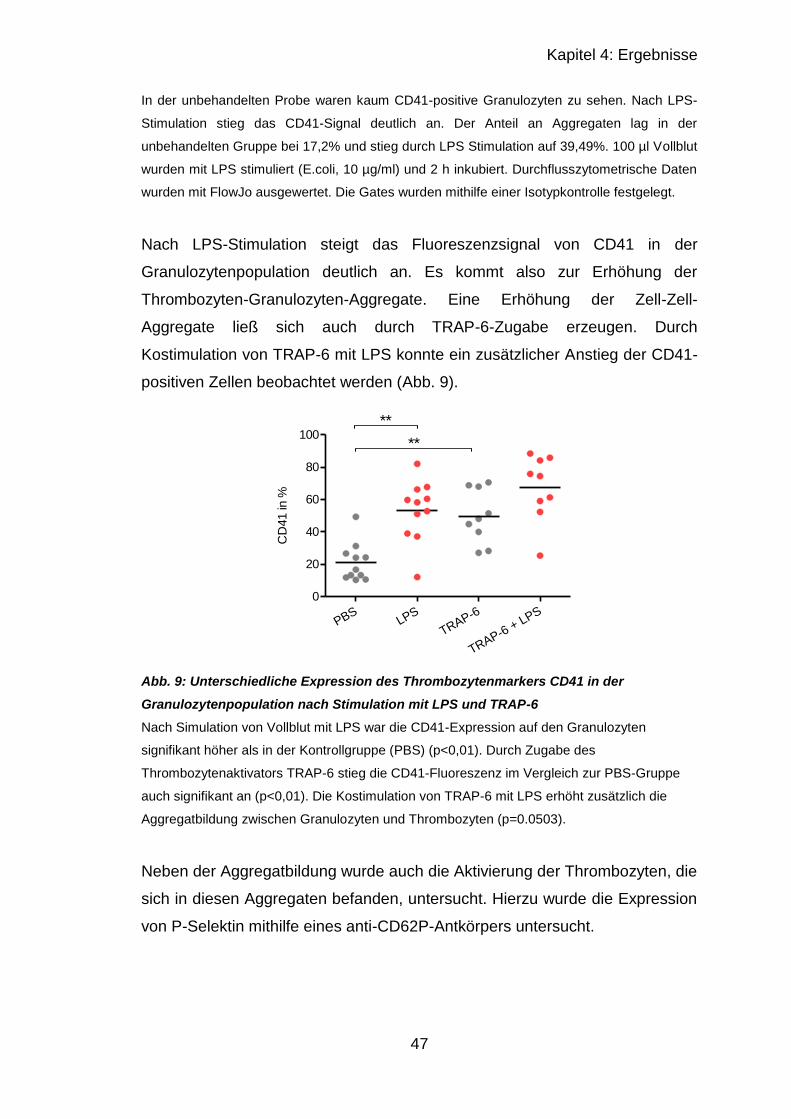
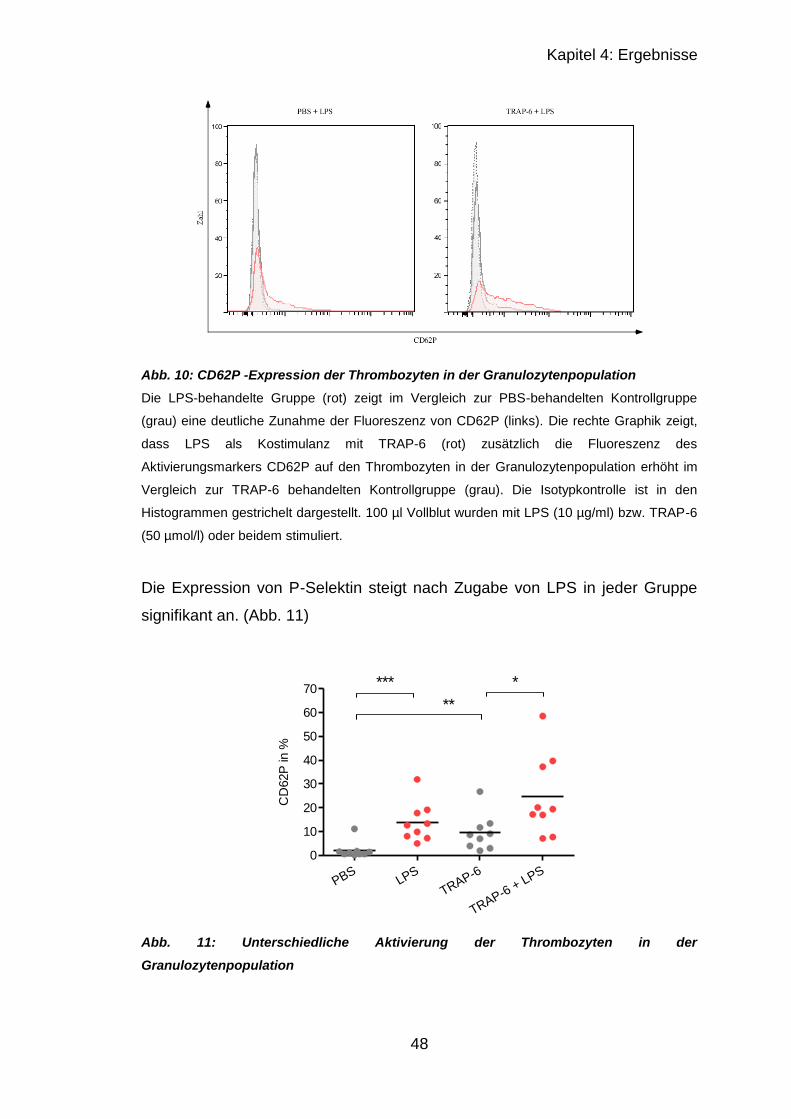



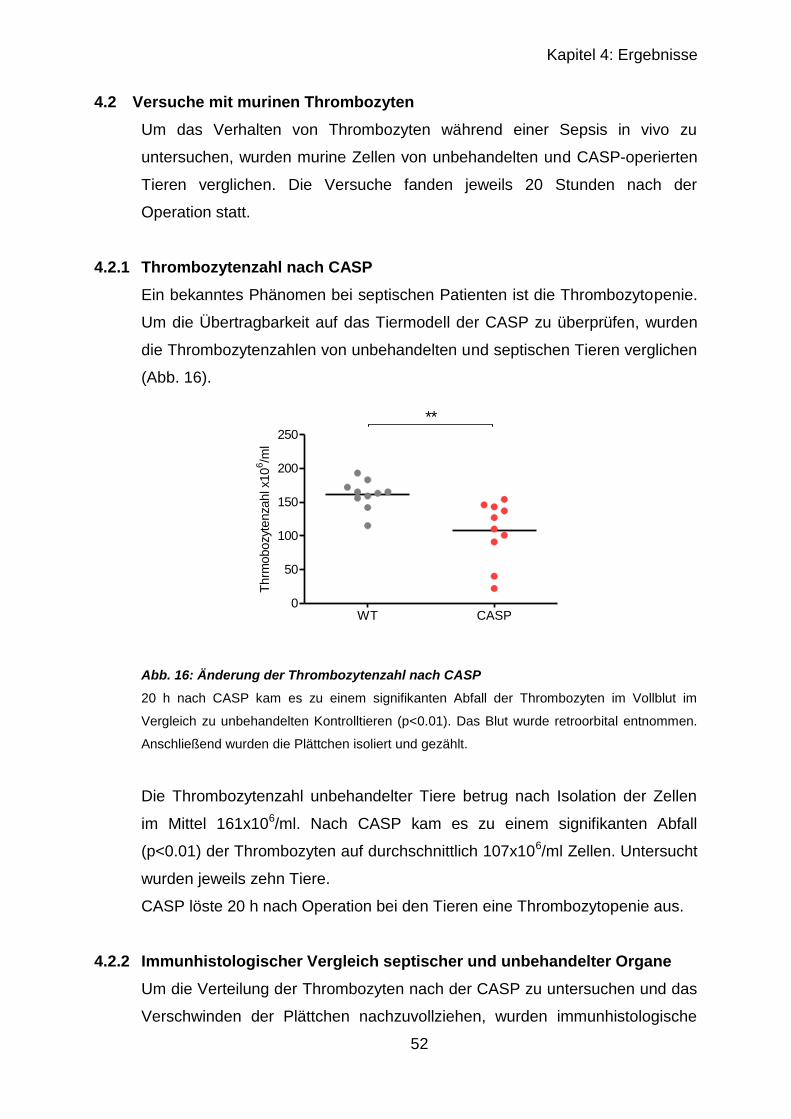
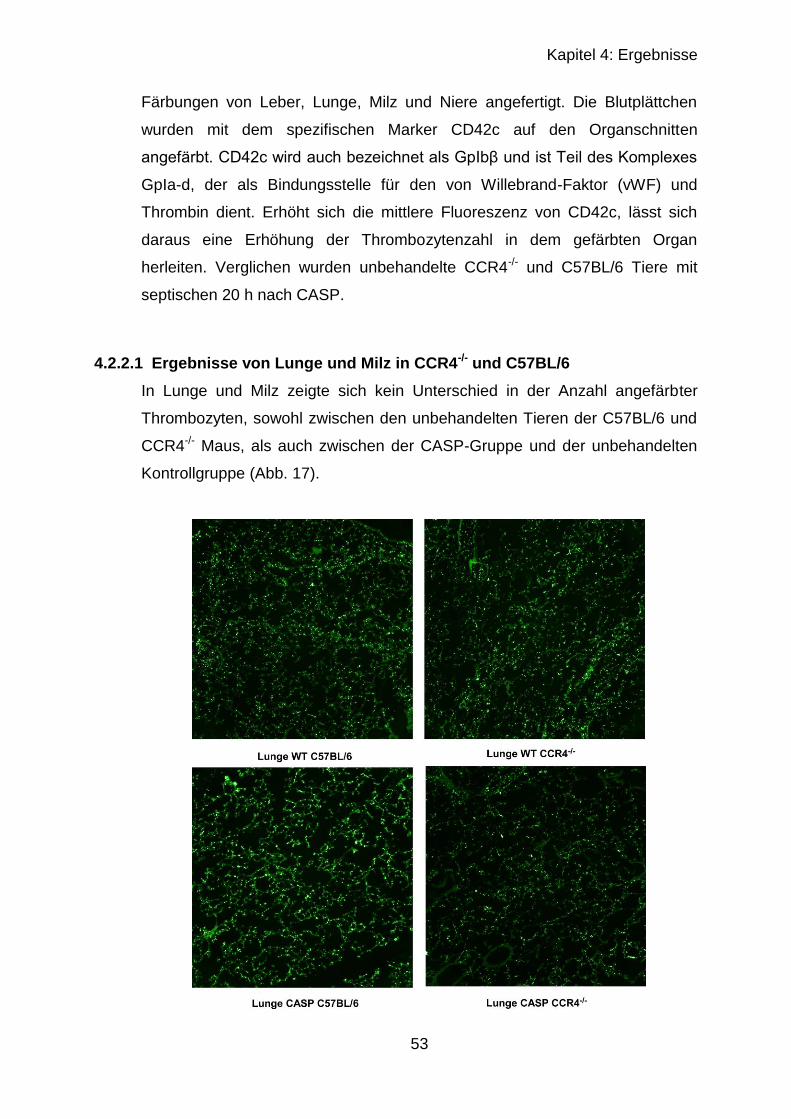
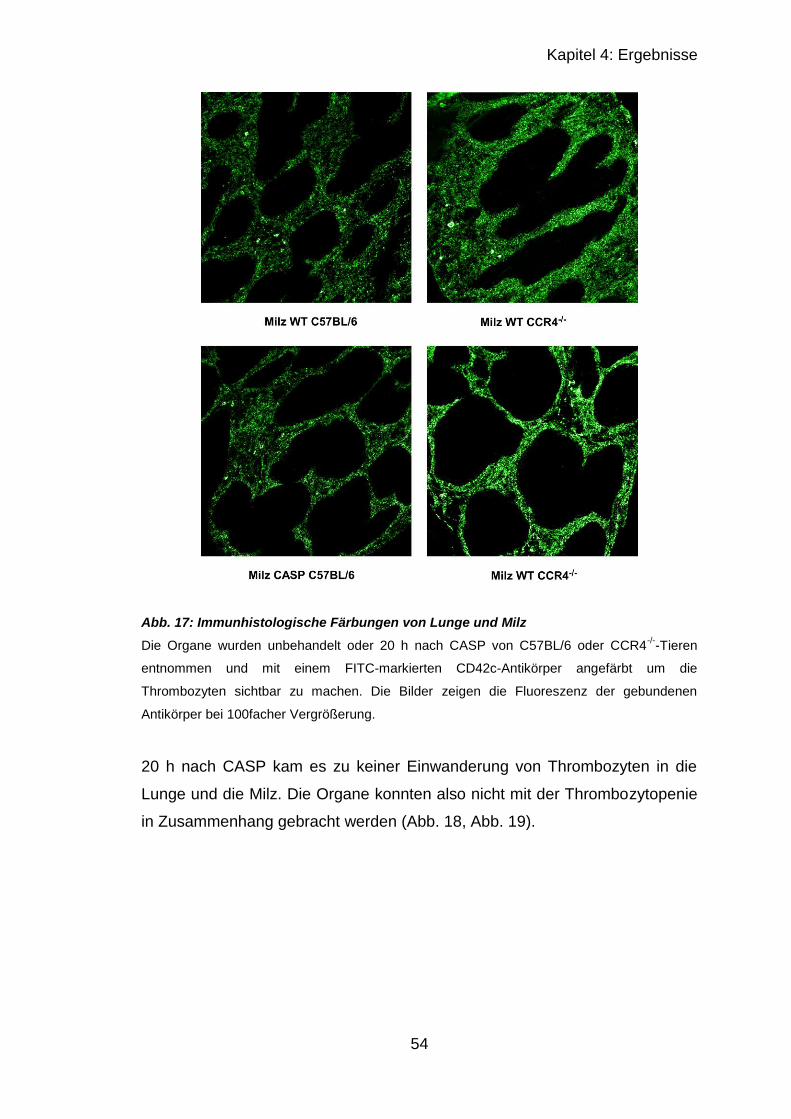
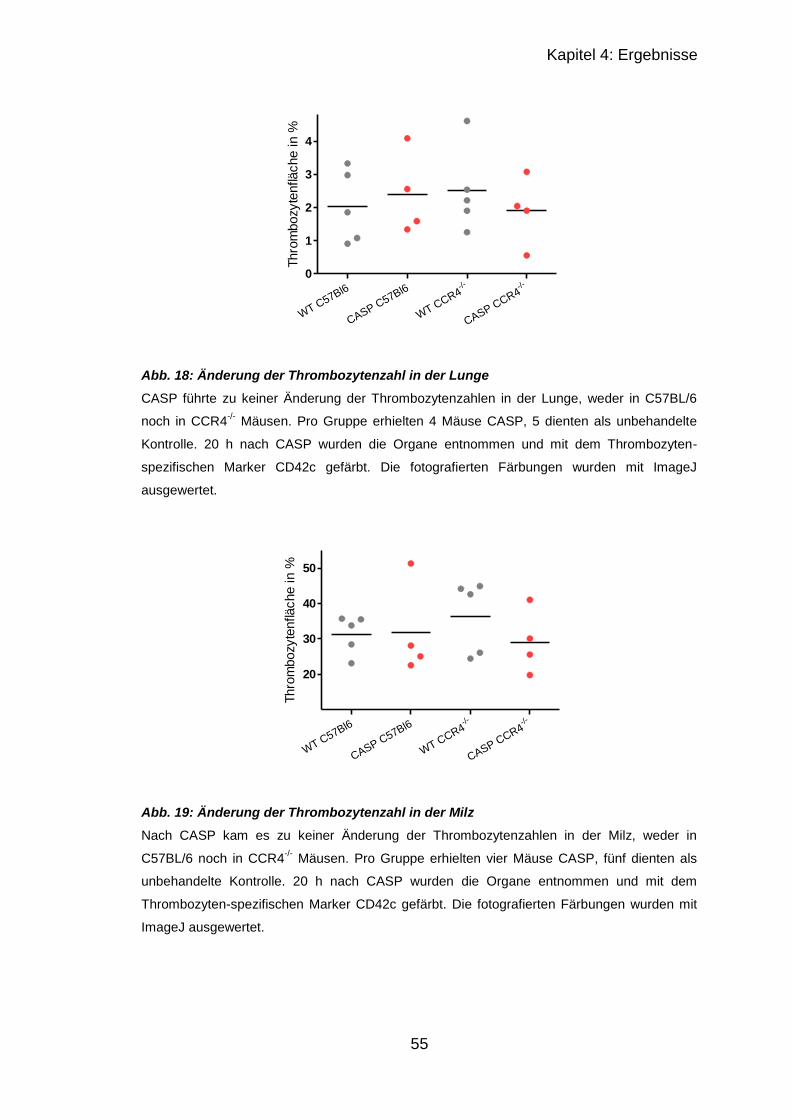
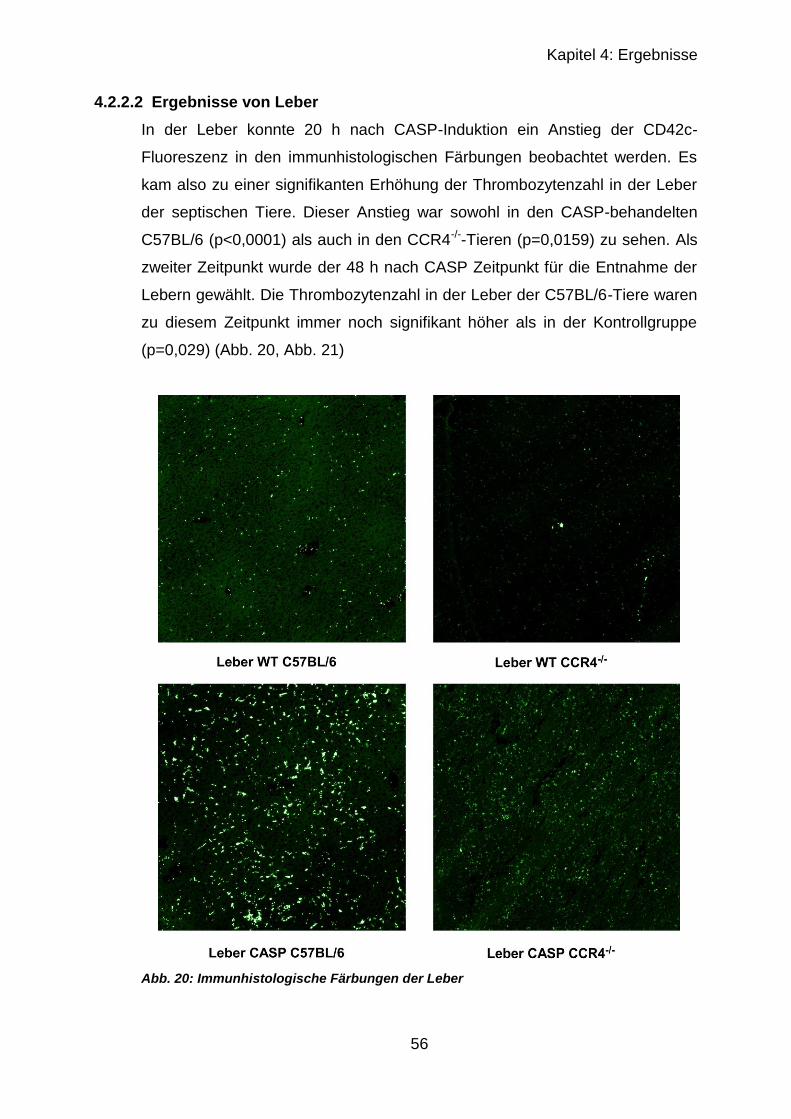
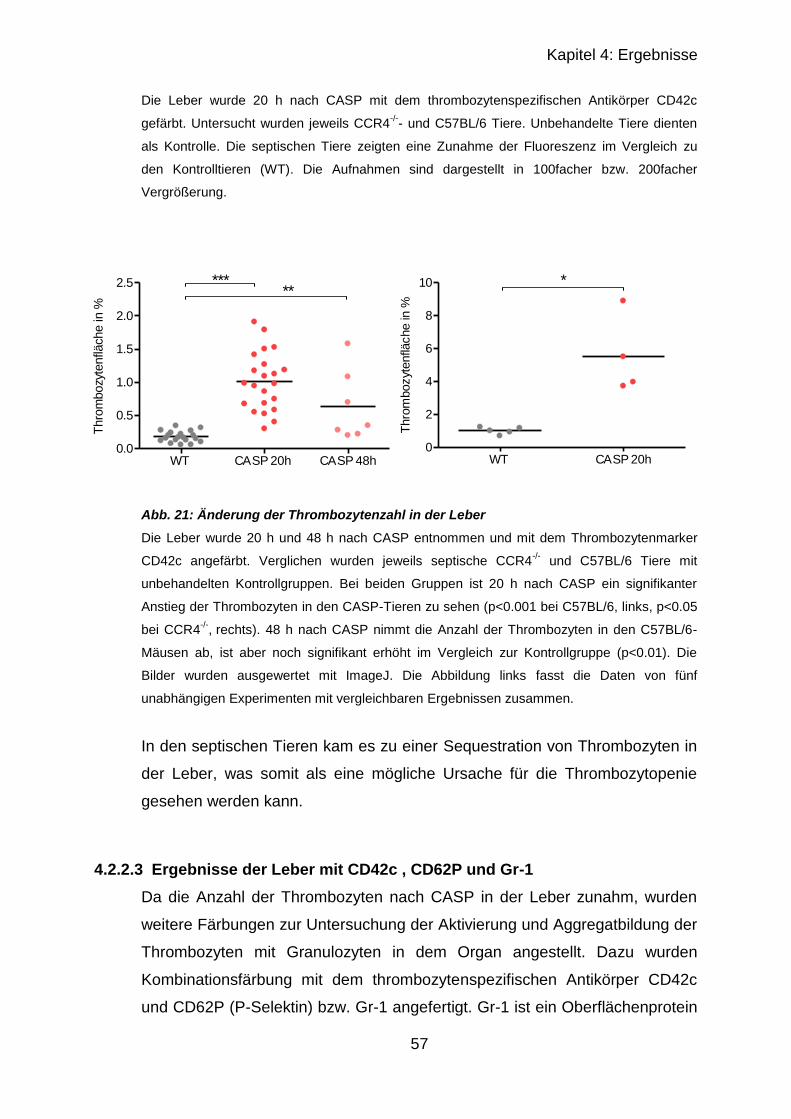


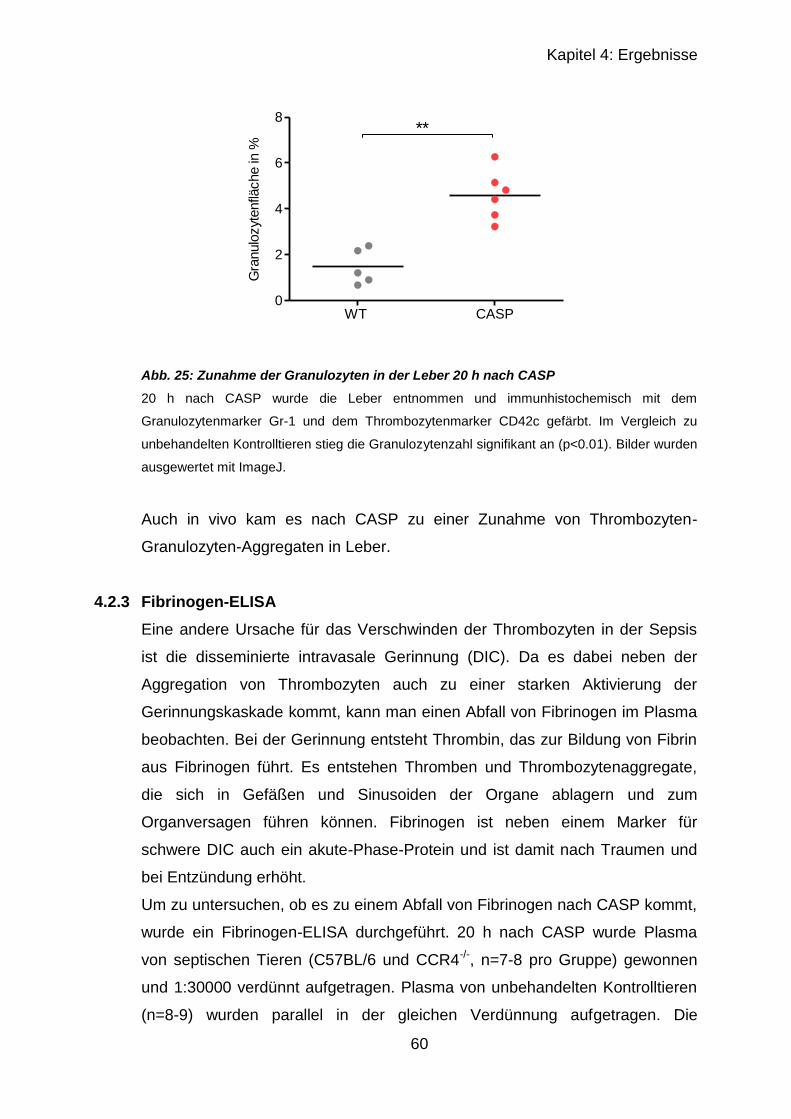
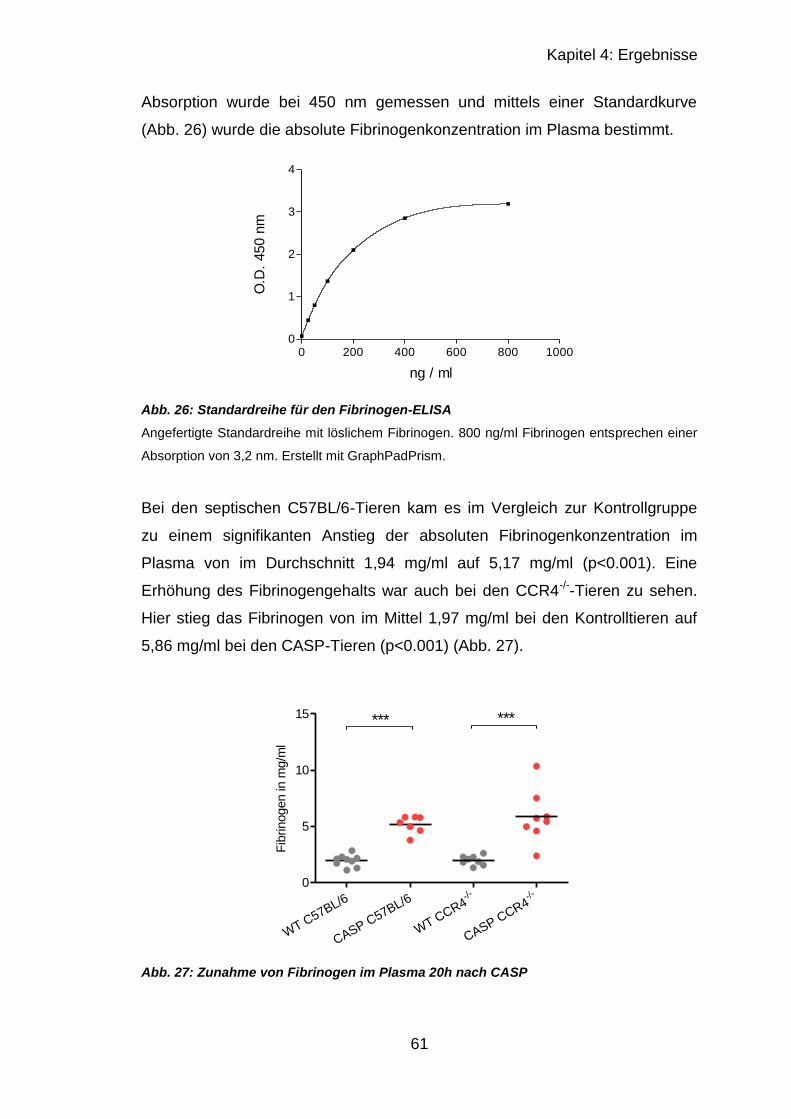

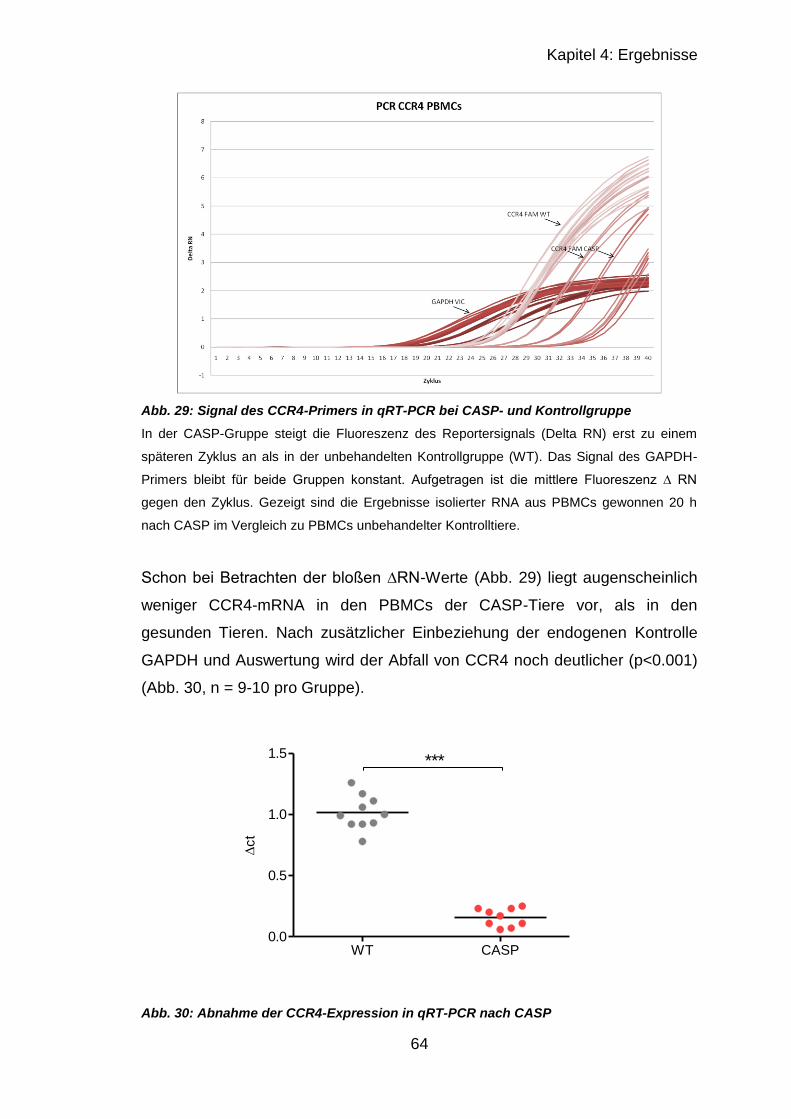










































![Perioperative Medikation [Schreibgesch tzt]new.st-elisabeth-hospital.de/fileadmin/Medizin/Anaesthesie/Referate... · Thrombozyten aggregationshemmer (TAH) • ASS verbessert die Prognose](https://static.unterlagen.site/doc/80x56/5d5990cc88c993d3668bb896/perioperative-medikation-schreibgesch-tztnewst-elisabeth-thrombozyten-aggregationshemmer.jpg)


