

Grenzen dynamischer Stabilität in komplexen biologischen Strukturen?
1. Netzwerke im Raum-Zeit-Kontinuum der belebten Welt
2. Zusammensetzung und Eigenschaften von Biozönosen („food webs“) –„Gesetzmäßigkeiten“
3. Erklärungen durch Modelle und experimentelle Beweise: Was macht sie stabil und (einige) instabil (chaotisch)?
4. Schlussfolgerungen 26 May, 9m

Aus Joyce &Palsson Nature Reviews 7 (2006)
„Informationswechsel“
Zelle: Metabolische Netzwerke - Stoffwechsel
1. Netzwerke im Raum-Zeit-Kontinuum der belebten Welt
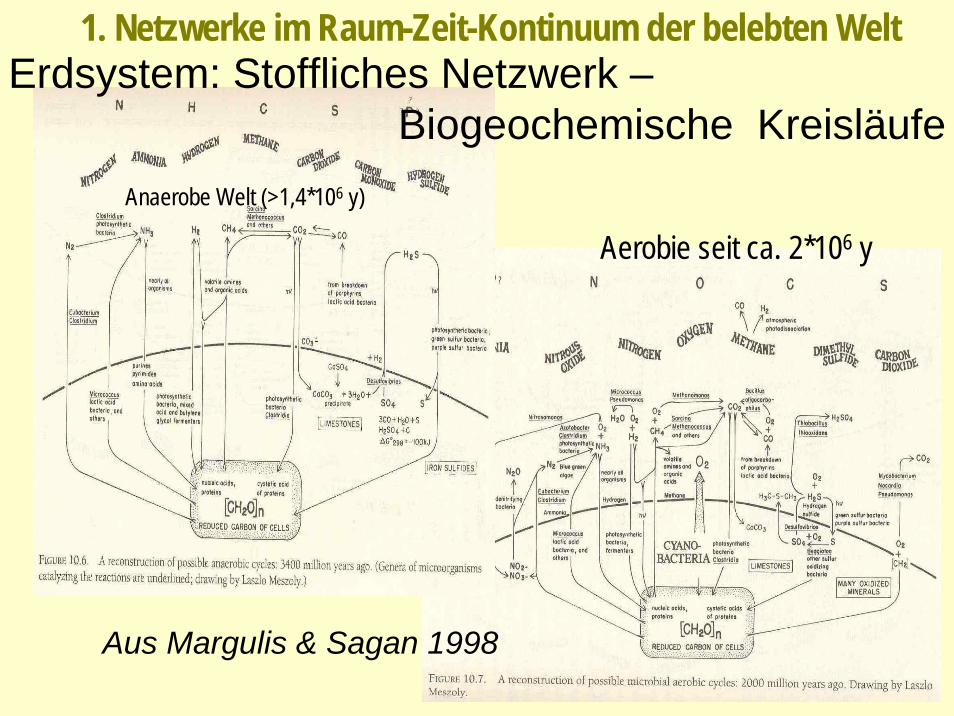
Aus Margulis & Sagan 1998
Erdsystem: Stoffliches Netzwerk –Biogeochemische Kreisläufe
Anaerobe Welt (>1,4*106 y)
1. Netzwerke im Raum-Zeit-Kontinuum der belebten Welt
Aerobie seit ca. 2*106 y
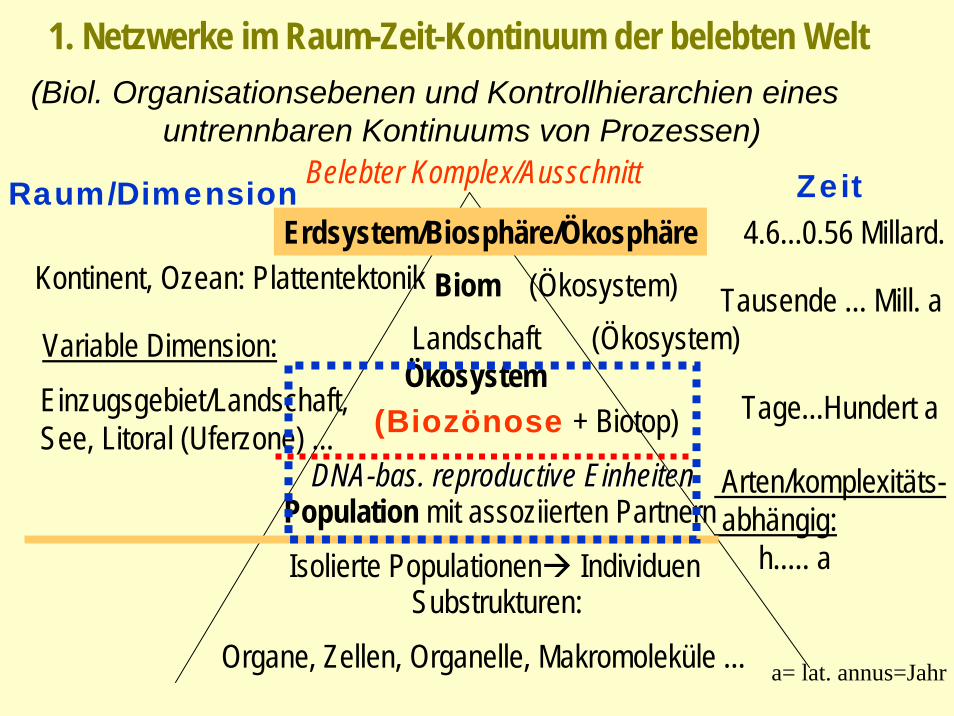
(Biol. Organisationsebenen und Kontrollhierarchien eines untrennbaren Kontinuums von Prozessen)
Raum/Dimension ZeitBelebter Komplex/Ausschnitt
Erdsystem/Biosphäre/ÖkosphäreBiom (Ökosystem)
4.6...0.56 Millard.
Landschaft (Ökosystem)Tausende ... Mill. a
Kontinent, Ozean: Plattentektonik
Ökosystem(Biozönose + Biotop)Einzugsgebiet/Landschaft,
See, Litoral (Uferzone) ...Tage...Hundert a
Variable Dimension:
DNADNA--basbas. . reproductivereproductive EinheitenEinheitenPopulation mit assoziierten Partnern
Arten/komplexitäts-abhängig:
h..... aIsolierte Populationen IndividuenSubstrukturen:
Organe, Zellen, Organelle, Makromoleküle ... a= lat. annus=Jahr
1. Netzwerke im Raum-Zeit-Kontinuum der belebten Welt
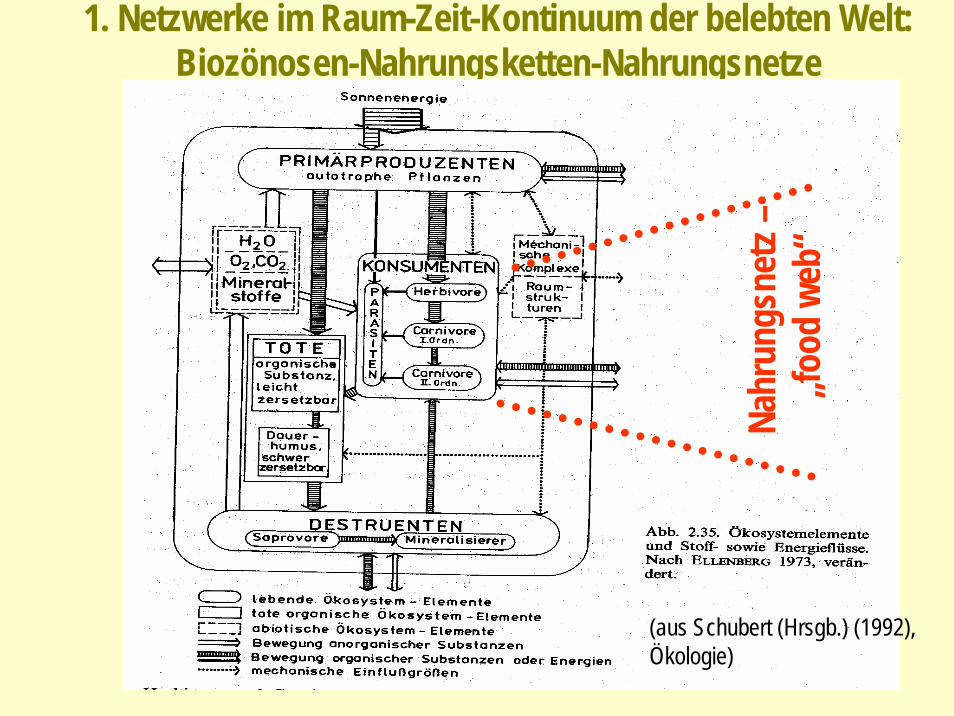
1. Netzwerke im Raum-Zeit-Kontinuum der belebten Welt: Biozönosen-Nahrungsketten-Nahrungsnetze
(aus Schubert (Hrsgb.) (1992), Ökologie)
Nahr
ungs
netz
–„fo
odwe
b“
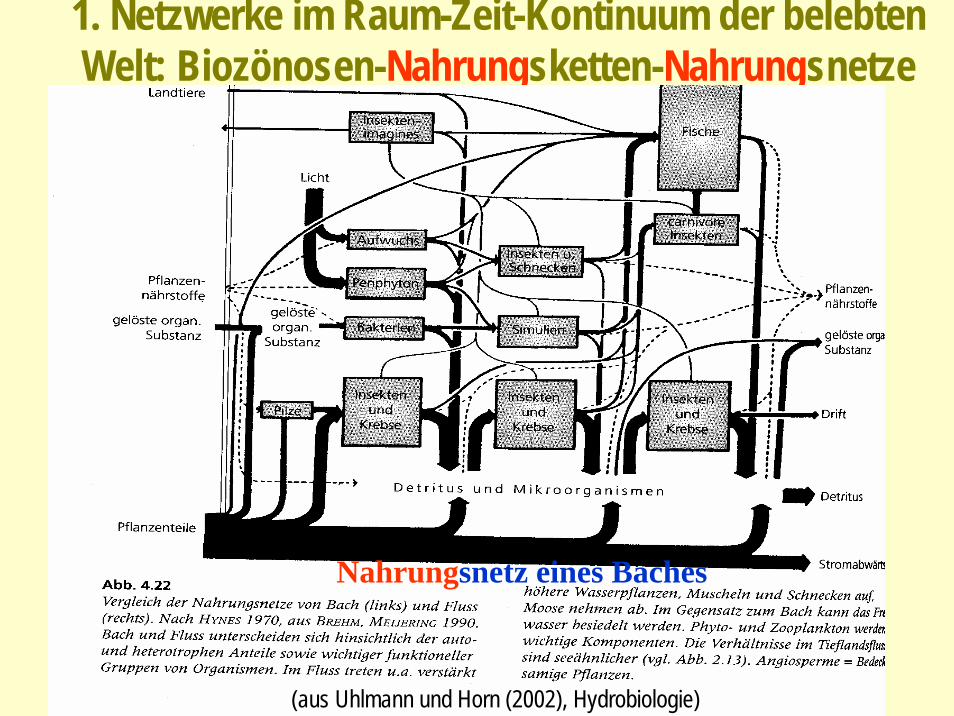
1. Netzwerke im Raum-Zeit-Kontinuum der belebten Welt: Biozönosen-Nahrungsketten-Nahrungsnetze
(aus Uhlmann und Horn (2002), Hydrobiologie)
Nahrungsnetz eines Baches

1. Netzwerke im Raum-Zeit-Kontinuum der belebten Welt
Das Konzept der (Lebens-)Gemeinschaft (Biozönose )→ Integration über zwischenartliche Wechselwirkungen
(Konkurrenz/Koexistenz, Mutualismen/Symbiosen) über hierarchische Kopplung und Interaktionen mit dem Lebensraum (Biotop) als wiederkehrende Muster(pattern):
→ Abstraktion von Gemeinschaftsmodulen (HOLT `97) oder funktionelle Gruppen in Kompartimenten (LAWTON `81)= funktionelle und taxonomische Aggregation (Beispiele):- Produzent-Konsument- Räuber - Beute - Ressource - Gilden (Arten, die dieselbe Ressourcenklasse nutzen)
trophische Ebenen = Ernährungsbeziehungen→ Starke Stofflich-energetische (und informelle) Kopplungen
(„Energie- und Stofffluss-Kanäle“) in Nahrungsnetzen mit Rückkopplungen und abgrenzbarer „Selbstregulation“
Biozönosen-Nahrungsketten-Nahrungsnetze
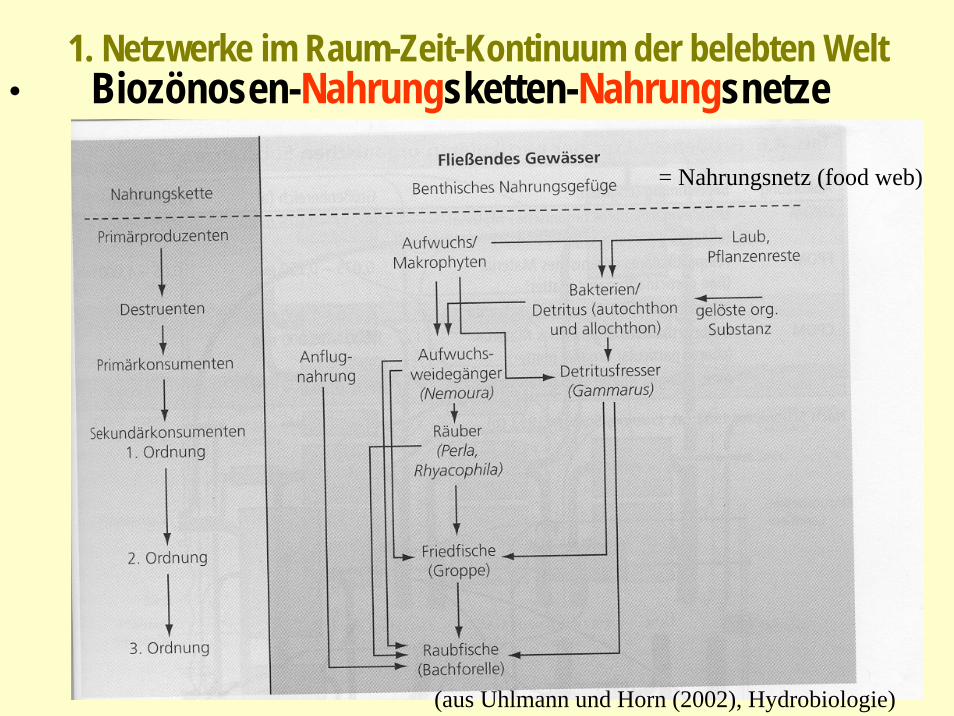
1. Netzwerke im Raum-Zeit-Kontinuum der belebten Welt• Biozönosen-Nahrungsketten-Nahrungsnetze
= Nahrungsnetz (food web)
(aus Uhlmann und Horn (2002), Hydrobiologie)

1. Netzwerke im Raum-Zeit-Kontinuum der belebten Welt
• Kompartimentierung: Stoffwechselkopplung• „Nahrungsketten“:Trophische Pyramide/Troph.Kaskade• Nahrungsnetz („food web“): „wer frißt wen?“:- emprische Netze (trophic links): Energie-/Stoff-Fluss- Wahrscheinliche Netze: Verbindungen auf wiederholt bestätigten Beziehungen (Pflanze-Weidegänger, Räuber- usw.)
- Imaginäre Netze: aus Artenlisten des Standortes abgeleitetAnalytische (empirische) Bestätigung auf der Basis der Stoff- und
Energieinhalte und der Änderungsraten zwischen den Kompartimenten
Energetisch-stoffliche Vernetzung (Trophodynamik)(Griechisch: Trophisch heißt von der Ernährung her betrachtet)

2. Zusammensetzung und Eigenschaften von „food webs“ -
Merkmale – „Gesetzmäßigkeiten“ (Regeln)Abstrakte Merkmale komplexer Interaktionen:- Verbindungen (graphisch: Linien) zwischen Konsument und
konsumierter Art(engruppe)- „Source webs, sinks webs, community webs“- Unidirektional (binäre Relation: Artenpaare) & direktional Interaktionen
(„links“ L) (Netto-Wirkung einer Art auf andere ohne intraspezifischeIteraktionen )
- Konnektivität (Connectance c) beschreibt wie viele Interakt. vorhanden sind: c = L/(S(S-1)/2) ... Wahrscheinlichkeit für ein zufällig gewähltes Artenpaar eine Wirkung auf andere zu entfalten (May 1973)
- Interaktionsdichte: Nahrungsbeziehungen (feeding links) je biologische Art (L/S)
Nach Cohen (1998) und Morin (2005)

Abstrakte Merkmale komplexer Interaktionen:- Verbindungen (graphisch: Linien) zwischen Konsument und
konsumierter Art(engruppe)- „Source webs, sinks webs, community webs“- Unidirektional (binäre Relation: Artenpaare) & direktional Interaktionen
(„links“ L) (Netto-Wirkung einer Art auf andere ohne intraspezifischeIteraktionen )
Nach Cohen (1998) und Morin (2005)
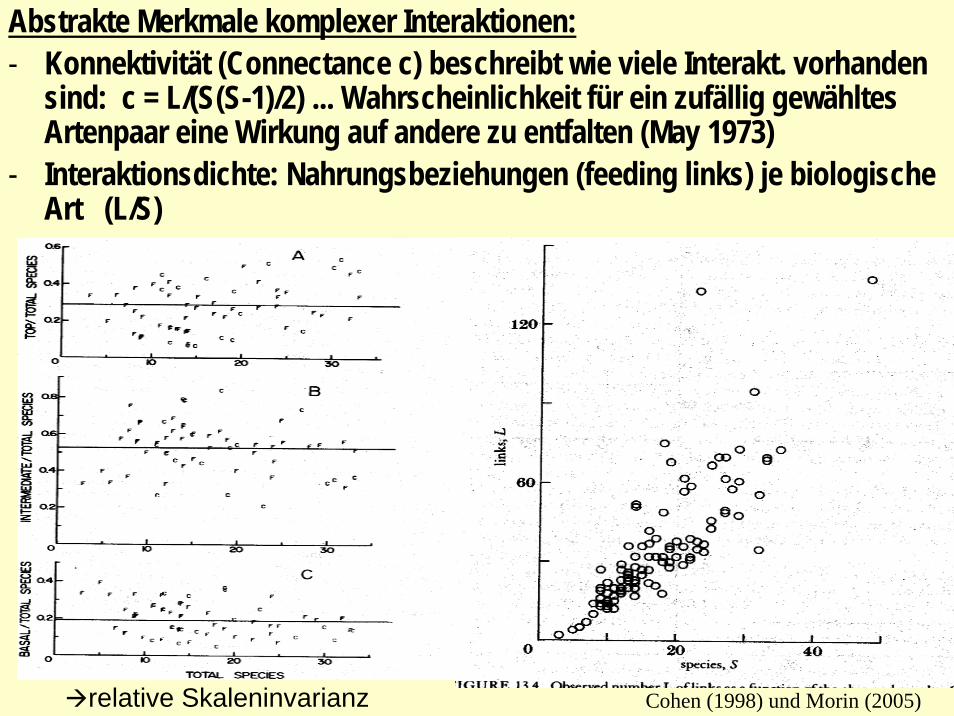
Abstrakte Merkmale komplexer Interaktionen:- Konnektivität (Connectance c) beschreibt wie viele Interakt. vorhanden
sind: c = L/(S(S-1)/2) ... Wahrscheinlichkeit für ein zufällig gewähltes Artenpaar eine Wirkung auf andere zu entfalten (May 1973)
- Interaktionsdichte: Nahrungsbeziehungen (feeding links) je biologische Art (L/S)
Cohen (1998) und Morin (2005)relative Skaleninvarianz

2. Zusammensetzung und Eigenschaften von „food webs“ -
Merkmale –„Gesetzmäßigkeiten“ (Regeln)Abstrakte Merkmale komplexer Interaktionen (2):- Kompartimentierung: beschreibt in welchem Umfang rel. isolierte
„subwebs“ enthalten sind - Trophische Ebene: Anzahl der Beziehungen +1 zwischen Basisarten
und interessierender Art- Omnivorie: tritt auf wenn Art andere konsumiert, die auf verschiedenen
trophischen Ebene Vorkommen oder in verschiedene Nahrungsketten (different chain omnivory)
- Schleifen: Arten konsumieren sich gegenseitig incl. Kannibalismus –selten und/oder wechselnd!
- Strenge „Kreisläufe“: Graphen für Überlappungen bei konsumierenden Arten (Räubern) in Bezug auf eine Beuteart
Nach Cohen (1998) und Morin (2005)

2. Zusammensetzung und Eigenschaften von „food webs“ -Eigenschaften
- Glieder (i.d.R. Räuber-Beute (Wirt-Parasit)-Beziehungen)selten zyklisch (d.h. gerichtet, ausgenommen Kannibalismus)verbunden über Knoten (direkt interagierende „Artenpaare“): Aggregation vieler Arten zu einer trophischen Kategorie (Knoten = „trophische Art“)
Kaskadenmodell- Ansammlung von (parallelen) Nahrungsketten (im Netz)- Nahrungsketten vertikal gerichtet (trophische Pyramiden)- Energietransfer zwischen Ketten ist nicht effizient - Nahrungsketten sind kurz (Gründe siehe unten: Stabilität)- Proportionen zwischen Spitzenkonsumenten, intermediären Arten und
Basis-Arten wenig variabel (Skaleninvarianz!)experimentelle Überprüfung (Tiere, Pflanzen: Mehrzeller)
Gewollte und ungewollte Steuerung von Nahrungsnetzen
)* empirisch ermittelt
aus Cohen (1989)
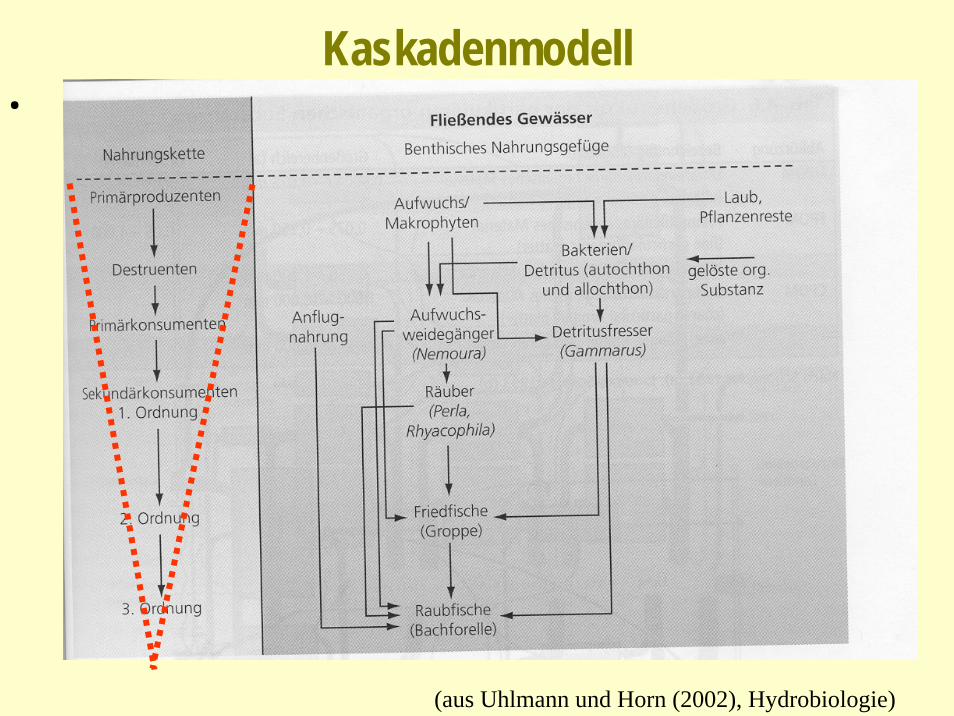
Kaskadenmodell.
(aus Uhlmann und Horn (2002), Hydrobiologie)
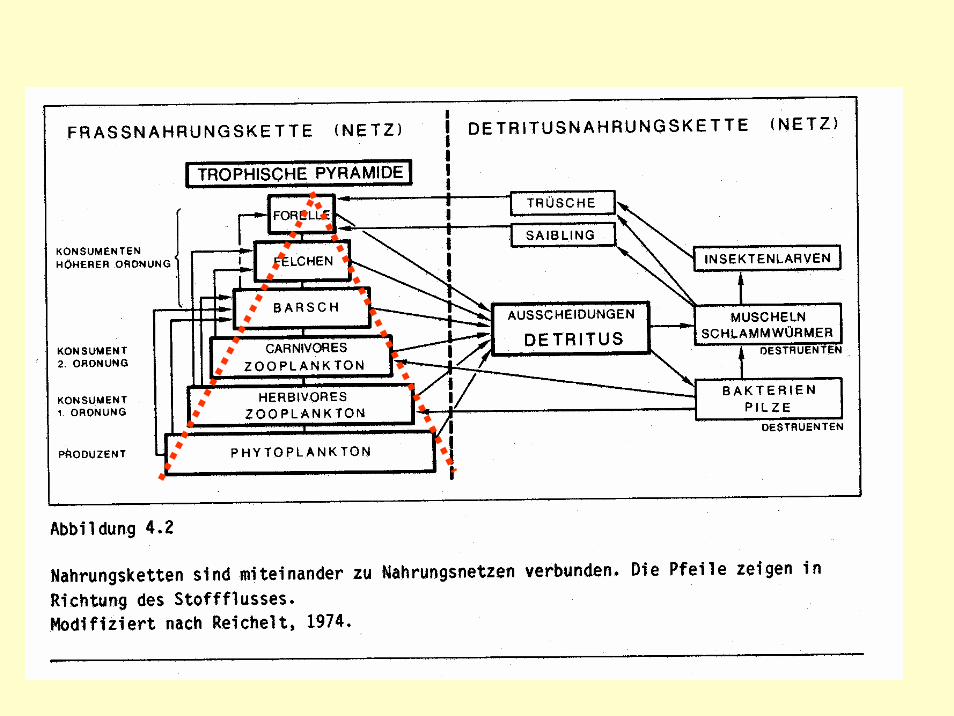
Abb.:

3. Erklärungen durch Modelle und experimentelle BeweiseErklärungen für Muster (Gesetzmäßigkeiten)
Basis:
I. Dynamische Räuber-Beute-Modelle nach Lotka-Volterra
II. Erweiterte Modelle mit > 2 Arten (May 1973, Pimm & Lawton1978)
III. Kaskadenmodell ohne spezifische Beziehung zwischen Populationsdynamik und Nahrungsnetzmuster (Cohen 1985)
aus Cohen (1989)Für ein System von n Arten sieht die Differntialgleichung für die Dynamik der Art i z.B. folgerndermaßen aus:
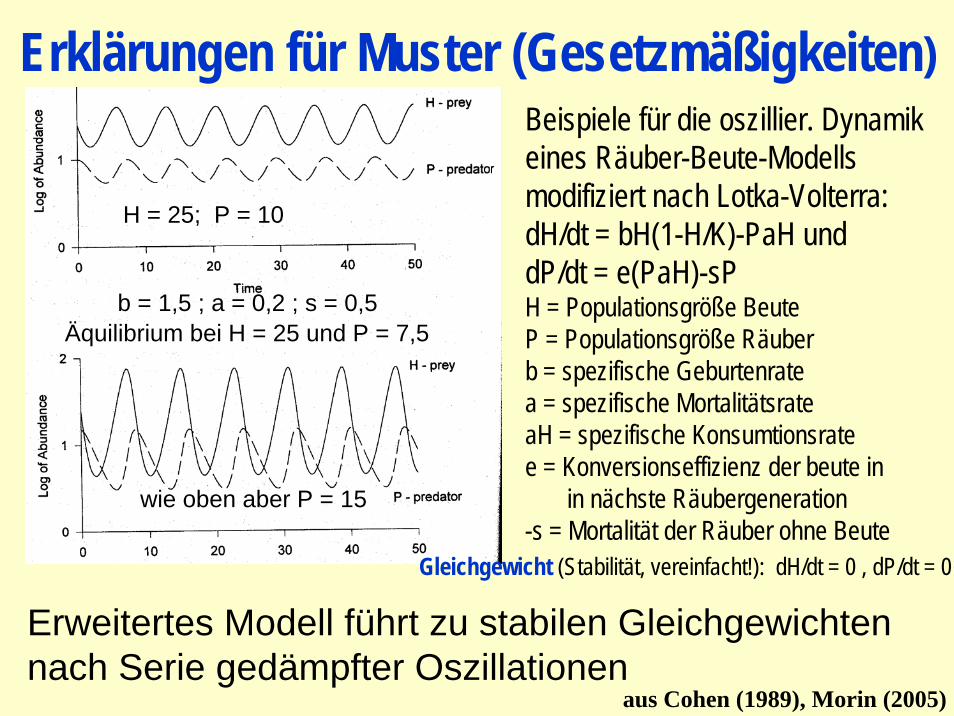
Erklärungen für Muster (Gesetzmäßigkeiten)
aus Cohen (1989), Morin (2005)
Beispiele für die oszillier. Dynamikeines Räuber-Beute-Modells modifiziert nach Lotka-Volterra:dH/dt = bH(1-H/K)-PaH unddP/dt = e(PaH)-sPH = Populationsgröße BeuteP = Populationsgröße Räuberb = spezifische Geburtenratea = spezifische MortalitätsrateaH = spezifische Konsumtionsratee = Konversionseffizienz der beute in
in nächste Räubergeneration-s = Mortalität der Räuber ohne Beute
Gleichgewicht (Stabilität, vereinfacht!): dH/dt = 0 , dP/dt = 0
b = 1,5 ; a = 0,2 ; s = 0,5
H = 25; P = 10
wie oben aber P = 15
Äquilibrium bei H = 25 und P = 7,5
Erweitertes Modell führt zu stabilen Gleichgewichten nach Serie gedämpfter Oszillationen
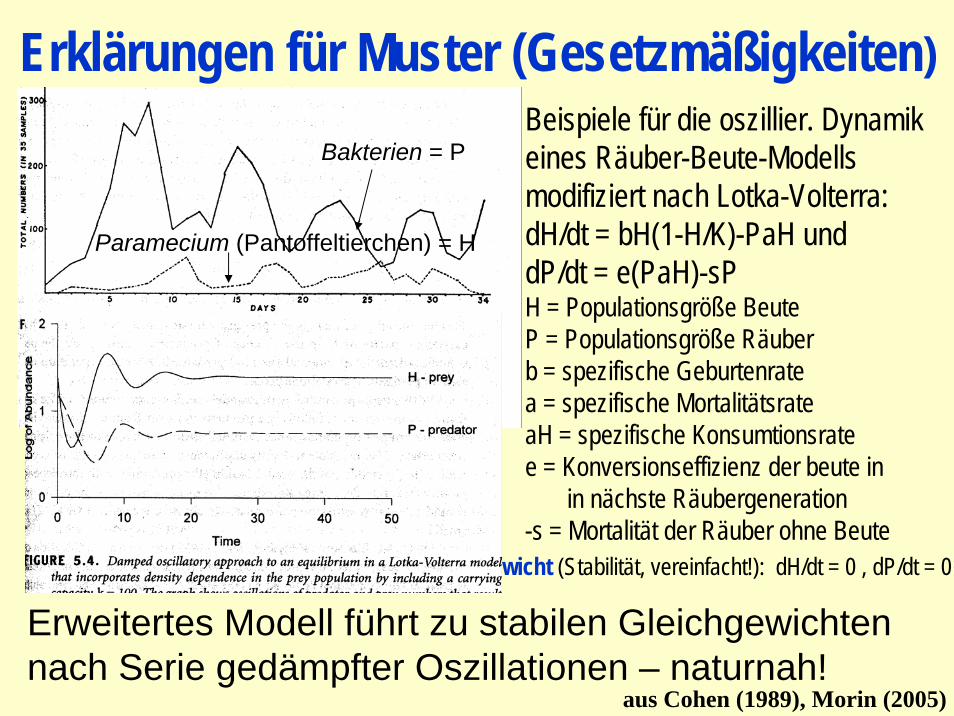
Erklärungen für Muster (Gesetzmäßigkeiten)
aus Cohen (1989), Morin (2005)
Beispiele für die oszillier. Dynamikeines Räuber-Beute-Modells modifiziert nach Lotka-Volterra:dH/dt = bH(1-H/K)-PaH unddP/dt = e(PaH)-sPH = Populationsgröße BeuteP = Populationsgröße Räuberb = spezifische Geburtenratea = spezifische MortalitätsrateaH = spezifische Konsumtionsratee = Konversionseffizienz der beute in
in nächste Räubergeneration-s = Mortalität der Räuber ohne Beute
Gleichgewicht (Stabilität, vereinfacht!): dH/dt = 0 , dP/dt = 0
Erweitertes Modell führt zu stabilen Gleichgewichten nach Serie gedämpfter Oszillationen – naturnah!
Bakterien = P
Paramecium (Pantoffeltierchen) = H

3. Erklärungen durch Modelle und experimentelle Beweise
Erklärungen für Muster (Gesetzmäßigkeiten)Basis:II. Erweiterte dynamische Räuber-Beute-Modelle
nach Lotka-Volterra mit > 2 Arten (May 1973, Pimm & Lawton 1977 ...)
dXi/dt = Xi(bi+Σai,jXj = Fi
Xi = Häufigkeit der Art i im System von n Artenbi = Netto-Wachstumsrate der Art iai,j = Wirkung (per capita effect) der Art i auf die Art j
( + / - / 0 )aus Cohen (1989)Für ein System von n Arten sieht die Differntialgleichung für die Dynamik der Art i z.B. folgerndermaßen aus:
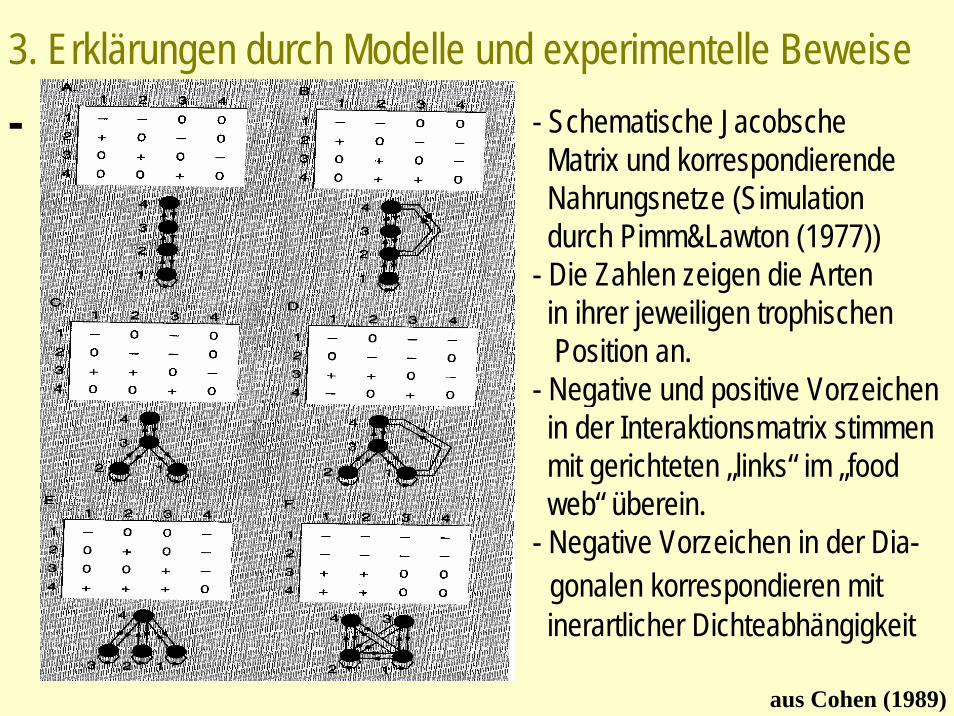
3. Erklärungen durch Modelle und experimentelle Beweise
-
aus Cohen (1989)
- Schematische Jacobsche Matrix und korrespondierende Nahrungsnetze (Simulation durch Pimm&Lawton (1977))
- Die Zahlen zeigen die Artenin ihrer jeweiligen trophischenPosition an.
- Negative und positive Vorzeichen in der Interaktionsmatrix stimmen mit gerichteten „links“ im „foodweb“ überein.
- Negative Vorzeichen in der Dia-gonalen korrespondieren mitinerartlicher Dichteabhängigkeit

3. Erklärungen durch Modelle und experimentelle Beweise
-
aus Cohen (1989)
- Häufigkeitsverteilung der Zeiten bis zum Eintritt des Äqulibriumsnach einer Störung (return time):
etwa 1/λ max = reziproker Wert des größten negativen Eigenwertes des Systems partieller Differential-gleichungen (jacobian matrix)
- Lange Nahrungsketten haben benötigen eine längere „return time“ (links oben A)
- Kurze gerichtete (direktionale) Nahrungsketten kehren schneller in‘s Gleichgewicht zurück (links E und C)
- Vernetzung erhöht die Variabilität
Rel
ativ
e A
ntei
le
Häufigkeitsverteilung (return time)

3. Erklärungen durch Modelle und experimentelle Beweise
Erklärungen für Muster (Gesetzmäßigkeiten)Ergebnisse der Modellierung:
1. Lange Nahrungsketten sind nicht stabil 2. Omnivorie destabilisiert Nahrungsketten
(...hat stärkeren Effekt als Länge der Kette!)3. Nahrungsnetze ohne genau (vor) determinierte Struktur
(random food webs) werden unstabiler, wenn ihre Komplexität, d.h. Artenvielfalt, Konnektivität, zunimmt(May, 1973, Pimm & Lawton 1978)
4. Zunehmende Komplexität kann Nahrungsnetze (sogar) destabilisieren
aus Cohen (1989), Morin (2005)
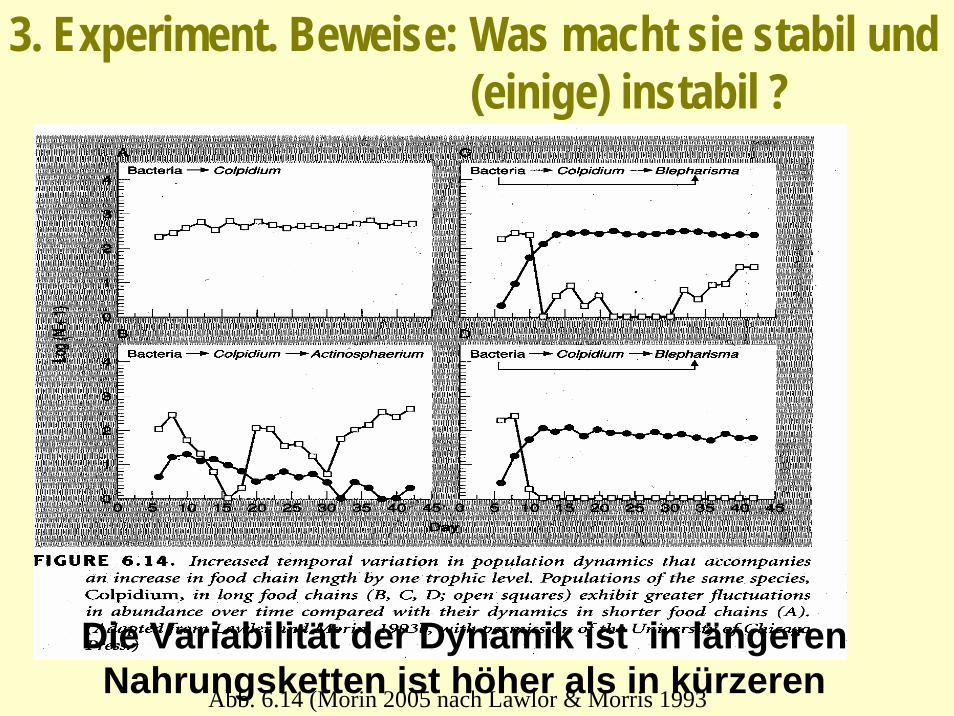
3. Experiment. Beweise: Was macht sie stabil und (einige) instabil ?
Abb. 6.14 (Morin 2005 nach Lawlor & Morris 1993
Die Variabilität der Dynamik ist in längeren Nahrungsketten ist höher als in kürzeren
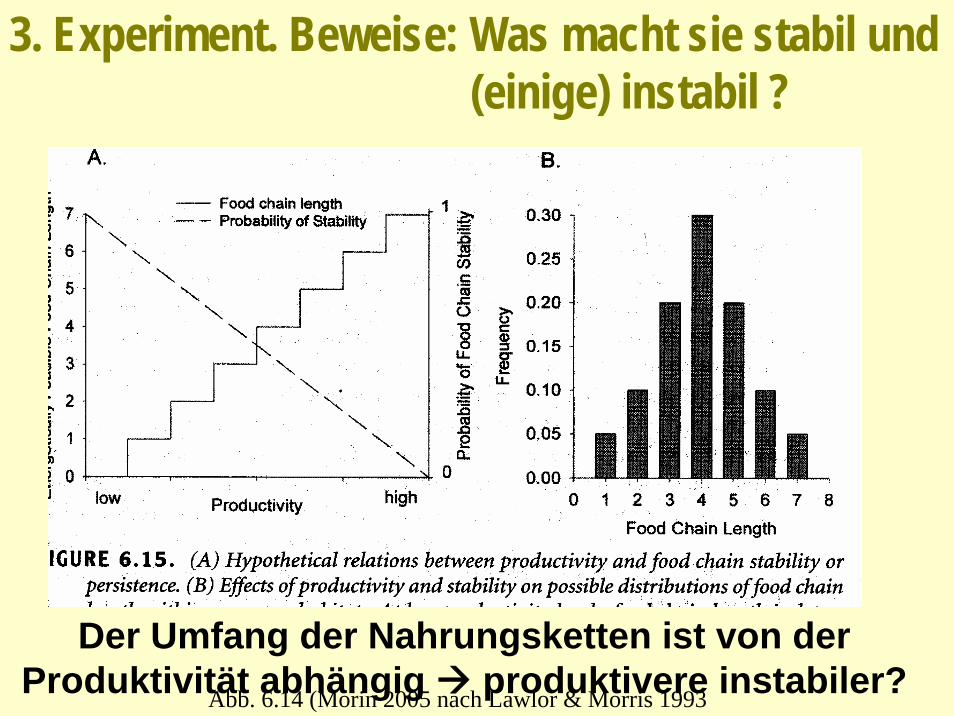
3. Experiment. Beweise: Was macht sie stabil und (einige) instabil ?
Abb. 6.14 (Morin 2005 nach Lawlor & Morris 1993
Der Umfang der Nahrungsketten ist von der Produktivität abhängig produktivere instabiler?

3. Experimentelle Beweise: Was macht sie stabil und (einige) instabil ?
3x Abb. 1 Becks et al. 2005
Innere Eigenschaften führen - in homogener Umwelt (!) - in einfachen „food webs“ zu Populations-schwankungen und deterministischen Chaos
Theorie: May (1974) unvorhersehbares VerhaltenExperimentelle Bestätigung:I. Zwei-Arten-Systeme (Rädertier-Alge: Fussmann
2000; Ciliat-Bakterie: Jost et al. 1973II. Drei-Arten-System (microbial food web!):
Becks, Hilker, Malchow, Jürgens & Arndt (2005) Posch,Mindl, Salcher, Sonntag, Vrba, Psenner(2006)

3. Experimentelle Beweise: Was macht sie stabil und (einige) instabil ?
3x Abb. 1 Becks et al. 2005
II. Drei-Arten-System (microbial food web!):(Heterotroph!)
Bedingungen:- Auswahl eines Räubers (Ciliat: Tetrahymena
pyriformis und zwei unterschiedlich bevorzugten Beutearten (Bakterien: stäbchenförmige Pedobacter(bevorzugt) und coccale Brevundimonas)
- Kultivierung im Fließgleichgewicht (Chemostat: konstanter Zu- und Abfluß=Durchflussrate
- Veränderung der Durchflussrate (Verweilzeit) als Bifurkationsparameter
Becks, Hilker, Malchow, Jürgens & Arndt in Nature 435 (2005)
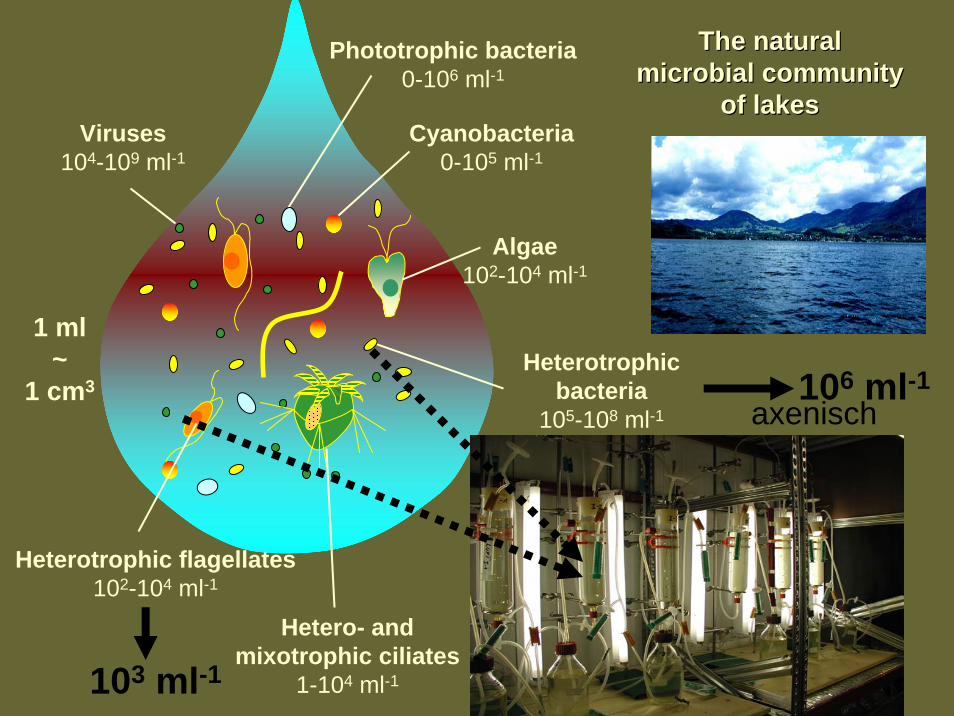
1 ml~
1 cm3
Heterotrophic flagellates102-104 ml-1
Heterotrophicbacteria
105-108 ml-1
Phototrophic bacteria0-106 ml-1
Viruses104-109 ml-1
Hetero- and mixotrophic ciliates
1-104 ml-1
Algae102-104 ml-1
Cyanobacteria0-105 ml-1
TheThe naturalnaturalmicrobialmicrobial communitycommunity
of of lakeslakes
103 ml-1
106 ml-1axenisch
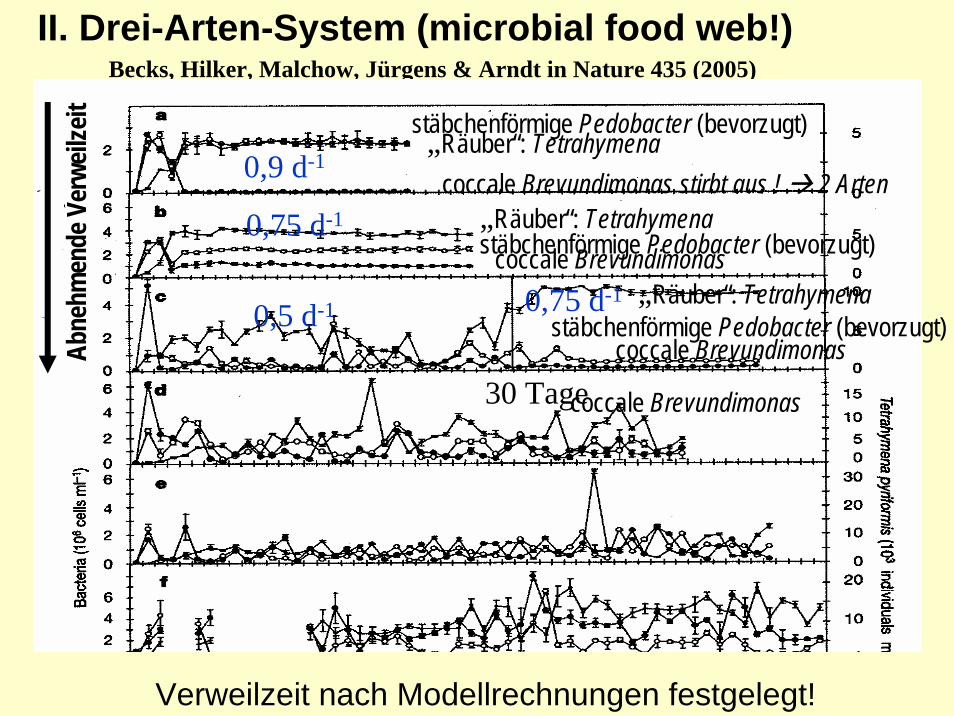
II. Drei-Arten-System (microbial food web!)Becks, Hilker, Malchow, Jürgens & Arndt in Nature 435 (2005)
stäbchenförmige Pedobacter (bevorzugt)
stäbchenförmige Pedobacter (bevorzugt)
stäbchenförmige Pedobacter (bevorzugt)
coccale Brevundimonas stirbt aus ! 2 Arten
coccale Brevundimonas
coccale Brevundimonas
coccale Brevundimonas
„Räuber“: Tetrahymena
„Räuber“: Tetrahymena
„Räuber“: Tetrahymena
Abne
hmen
de V
erwe
ilzeit
0,9 d-1
0,5 d-1
0,75 d-1
0,75 d-1
30 Tage
Verweilzeit nach Modellrechnungen festgelegt!
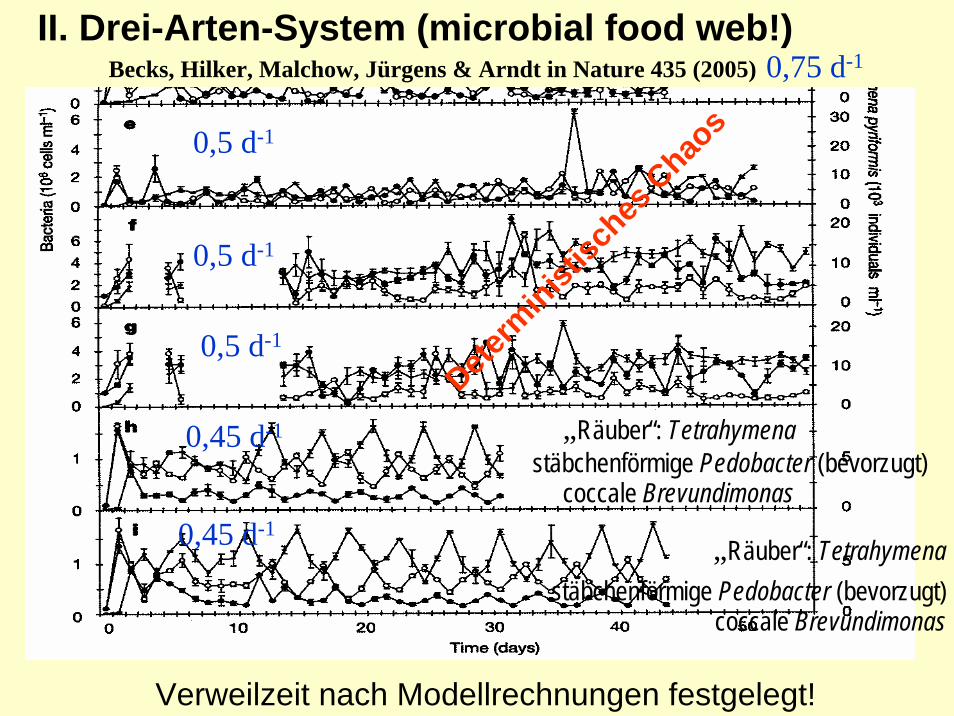
II. Drei-Arten-System (microbial food web!)Becks, Hilker, Malchow, Jürgens & Arndt in Nature 435 (2005)
stäbchenförmige Pedobacter (bevorzugt)
stäbchenförmige Pedobacter (bevorzugt)coccale Brevundimonas
coccale Brevundimonas
„Räuber“: Tetrahymena
„Räuber“: Tetrahymena
0,5 d-1
0,45 d-1
0,75 d-1
0,5 d-1
0,45 d-1
0,5 d-1
Determ
inistisc
hes C
haos
Verweilzeit nach Modellrechnungen festgelegt!

3. Experimentelle Beweise: Was macht sie stabil und (einige) instabil ?
II. Drei-Arten-System (microbial food web!)
Becks, Hilker, Malchow, Jürgens & Arndt in Nature 435 (2005)

Schlussfolgerungen
• Nahrungsketten sind kurz• Mit zunehmender Länge nimmt Interaktions-
stärke in der Regel deutlich ab • Sie sind kompartimentiert („geblockt“) und
in „sub webs“ strukturiert • Nahrungsnetze bezüglich der Interaktionen
stark begrenzt (begrenzt Komplex) • (deterministisches) Chaos – intrinsische
Stochastizität ist möglich (vgl. Umweltstochastizität)
unter streng deterministischen Randbedingungen
(bezüglich Stabilitätsförderung so selektiert)

Schlussfolgerungen (2)
• Aussagen sind nur für die jeweilige Komplexitätsebene gültig (vgl. Einleitung)
- Große (vielzellige) Tiere (Raum-Zeit-Dimension des Beobachters) - Mikrobielle - homogene - Nahrungsnetze (Chemostaten, Seen bzw. ihr durchmischtes Pelagial)
• Sie gelten für weitgehend geschlossene Systeme- Migration und Extinktion sind ausgeschlossen- Ressourcen werden „recycelt“ bzw. Energie nachgeliefert• Irreguläre Umweltveränderungen undGradienten sind ausgeschlossen (Umweltstochastizität)
• Sie beziehen sich nur bzw. berücksichtigen ausschließlich stofflich-energetische Nahrungsbeziehungen (s.u.)
informelle Interaktionen sind nicht berücksichtigt bzw. subsummiert
(bezüglich Prämissen)

Abb.:
Scheu et al. 2001)
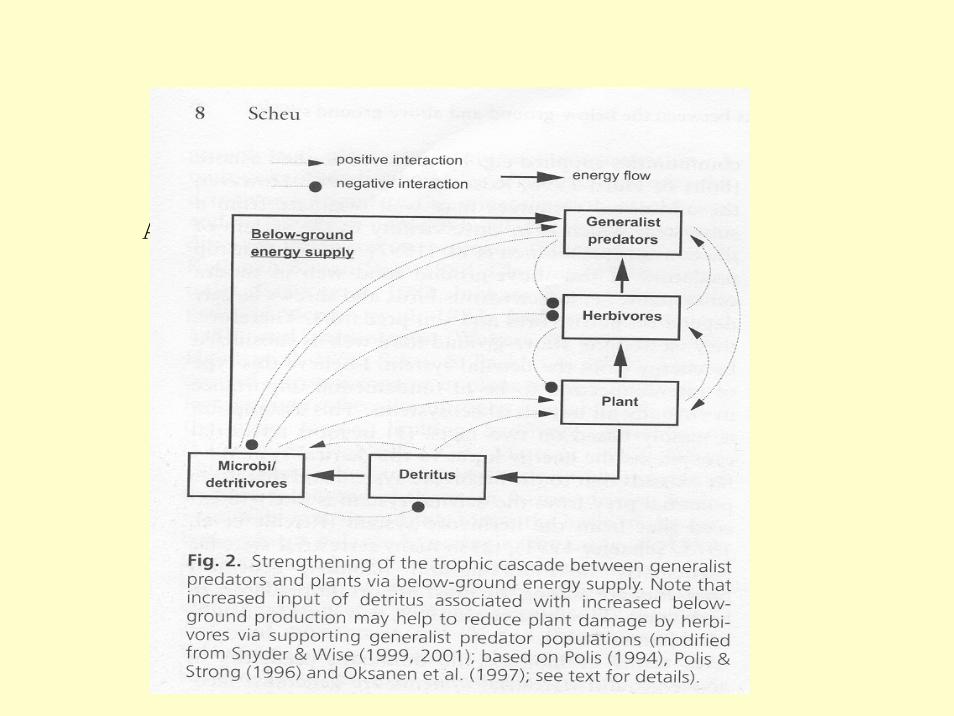
Abb.:
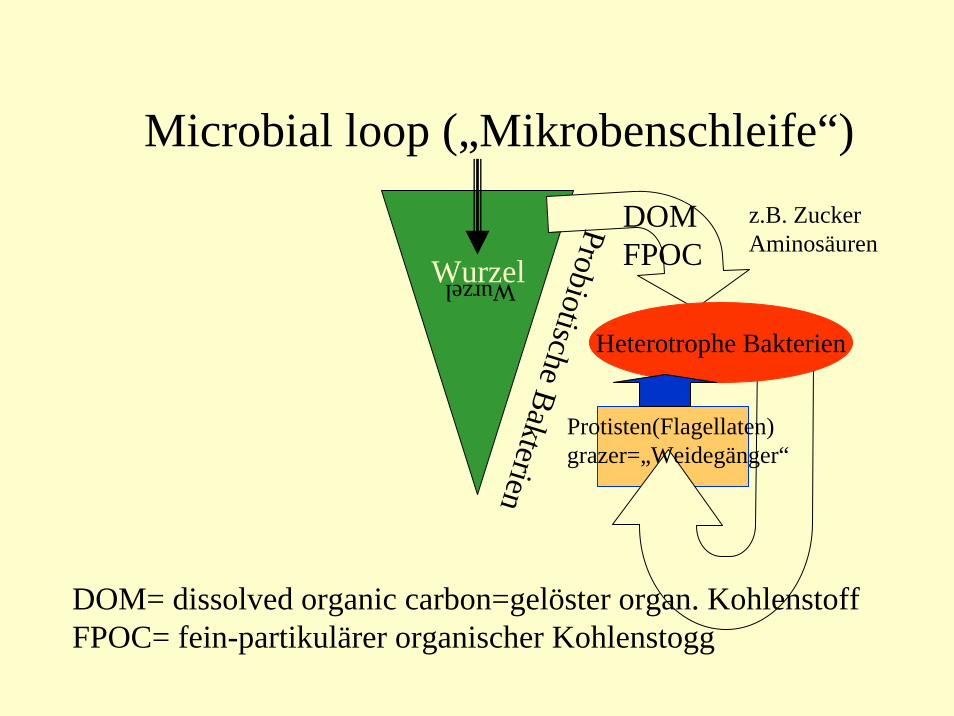
Microbial loop („Mikrobenschleife“)
Wurzel
DOMFPOCWurzel
DOM= dissolved organic carbon=gelöster organ. KohlenstoffFPOC= fein-partikulärer organischer Kohlenstogg
Heterotrophe Bakterien
Protisten(Flagellaten)grazer=„Weidegänger“
ProbiotischeBakterien
z.B. ZuckerAminosäuren
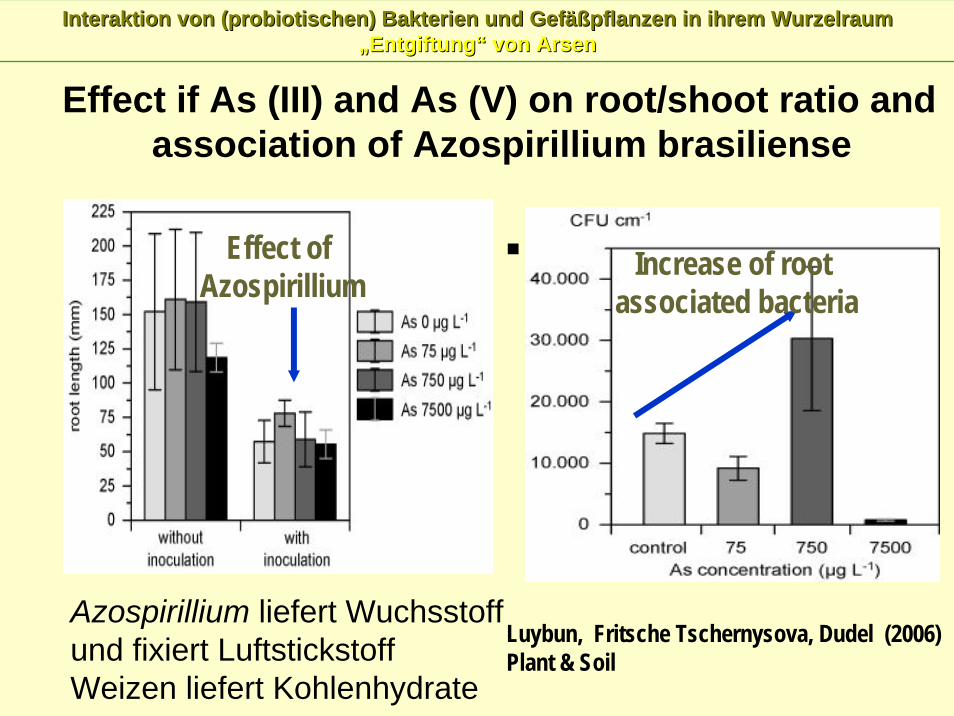
Interaktion von (Interaktion von (probiotischenprobiotischen) Bakterien und Gef) Bakterien und Gefäßäßpflanzen in ihrem Wurzelraum pflanzen in ihrem Wurzelraum „„EntgiftungEntgiftung““ von Arsenvon Arsen
Luybun, Fritsche Tschernysova, Dudel (2006)Plant & Soil
Effect of Azospirillium Increase of root
associated bacteria
Effect if As (III) and As (V) on root/shoot ratio and association of Azospirillium brasiliense
Azospirillium liefert Wuchsstoffund fixiert LuftstickstoffWeizen liefert Kohlenhydrate

Schlussfolgerungen (2)
• Aussagen gelten nur für die jeweilige Skalenebene(Mikroben, Tiere usw.) und sofern die Umwelt im dynamischen Gleichgewicht ist
• Ihre Gültigkeit für inhomogene Umwelten ist nicht bewiesen (System Pflanze-Rhizosphäre-Boden/org. Auflage) bzw. für „Subsysteme“ vergleichbarer Dimension gültig
• Aussagen besitzen daher nur eine sehr begrenzteVerallgemeinerungsfähigkeit
• Sie gelten nicht oder nur begrenzt für Phasengrenzenbzw. Grenzschichten (Ökotone, Biofilme usw.)
• Aussagen besitzen daher nur eine sehr begrenzteVerallgemeinerungsfähigkeit
(Gültigkeitsbereich der Aussagen)

Dank
an Hartmut Arndt und Mitarbeiter (Universität Köln)Thomas Posch und Mitarbeiter (Universität Innsbruck)
und an Sie für Ihre Aufmerksamkeit!
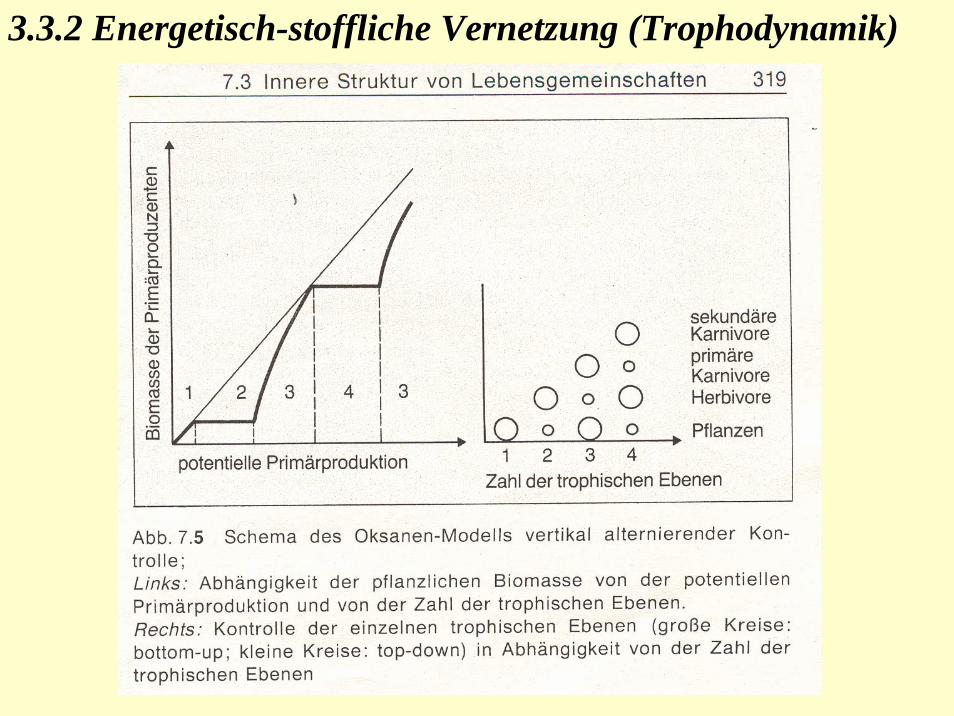
3.3.2 Energetisch-stoffliche Vernetzung (Trophodynamik)
Abb.:







