Page 1
1
Aus der
Medizinischen Klinik I
Marienhospital Herne
- Universitätsklinik -
Der Ruhr-Universität-Bochum
Kommissarischer Direktor: Prof. Dr. R. Voigtmann
____________________________________________________________________
Wirkung von Eisen-III-natrium-gluconat-Komplex auf die
Produktion von
reaktiven Sauerstoffradikalen bei Patienten mit terminaler
Niereninsuffizienz
Inaugural-Dissertation
zur
Erlangung des Doktorgrades der Medizin
einer
Hohen Medizinischen Fakultät
der Ruhr-Universität Bochum
vorgelegt von
Anna Matuszczyk
aus Ruda/Kattowitz
2001
Page 2
2
Dekan: Prof. Dr. med. G. Muhr
Referent: Prof. Dr. med. M. Tepel
Koreferent: Prof. Dr. med. Wolf – H. Schmiegel
Tag der mündlichen Prüfung:
12.11.2002
Page 3
3
Meiner lieben Familie gewidmet
Page 4
4
INHALT
Seite 1. Einleitung 6
2. Problemstellung 12
3. Materialien 13
3.1 Materialien und Reagenzien 13
3.2 Geräte 14
3.3 Lösungen 14
4. Methoden 15
4.1 Patientencharakterisierung 15
4.2 Prinzip der Fluoreszenz-Spektrophotometrie 18
4.3 Präparation von mononukleären Leukozyten 18
4.4 Messung von mononukleären Leukozyten 19
4.5 Messung von Oxidantien und Antioxidantien 19
4.6 Messung von Vanadate, Disulfiram, DOG und
Eisen-III-natrium-gluconat-Komplex 21
4.7 Messung mit FeCl3, FeCl2 und Trolox 21
4.8 Versuche mit dem Eisenindikator phen green 22
4.9 Kontrollversuche in Abwesenheit von
mononukleären Leukozyten 23
4.10 Statistik 24
5. Ergebnisse 25
5.1 Wirkung von Oxidantien und Antioxidantien 25
5.2 Wirkung von Vanadate, Disulfiram, DOG und
Eisen-III-natrium-gluconat-komplex 35
5.3 Wirkung von FeCl3, FeCl2 und Trolox 38
5.4 Versuche mit dem Eisenindikator phen green 41
5.5 Kontrollversuche in Abwesenheit von
mononukleären Leukozyten 45
6. Diskussion 47
7. Zusammenfassung 52
8. Literaturverzeichnis 56
9. Danksagungen 69
10. Lebenslauf 74
Page 5
5
ABKÜRZUNGEN
Ferrlecit = Eisen-III-natrium-gluconat-Komplex
SOD = Superoxiddismutase
Ebselen = 2-Phenyl-1,2-benzisoselenazol-3(2H)-one
NaN3 = Natriumazid
PMA = Phorbol-12-myristat-13-acetat
FeCl3 = Dreiwertiges Eisenchlorid
FeCl2 = Zweiwertiges Eisenchlorid
EXP- 3174 = Losartan
DOG = 1,2 Dioctanoyl-sn-glycerol
Trolox = 6-Hydroxy-2,5,7,8-tetramethylchroman-2-carboxylsäure
DCFDA = 2`,7`-Dichlorofluoreszindiacetat
DCF = 2`,7- Dichlorofluoreszin
NAD+ = Nicotinamidadenindinucleotid ( oxidierte Form )
NADH = Nicotinamidadenindinucleotid ( reduzierte Form )
NADP+ = Nicotinamidadenindinucleotidphosphat
( oxidierte Form )
NADPH = Nicotinadenindinucleotidphosphat ( reduzierte Form )
EPO = Erythropoetin
HBSS = „Hank´s balanced salt solution“
Page 6
6
1. EINLEITUNG
Das Überleben aerober Zellsysteme wird durch die Reduktion des molekularen
Sauerstoffs zu Wasser während der Atmungskette in den Mitochondrien gesichert.
Molekularer Sauerstoff, O2, kann jedoch die Bildung von Sauerstoffradikalen mit
einem hohen toxischen Potential verursachen.
Zu ihnen gehören unter anderem der Singulett-Sauerstoff, das Superoxid-Radikal-
Anion, das Superoxidradikal, das Hydroxyl-Radikal und Wasserstoffperoxid.
Während der Singulett-Sauerstoff, ?gO2, sich in einem besonderen elektronischen
Anregungszustand befindet, der z.B auch bei Einwirkung von hochenergetischen
Strahlen auf O2 entsteht, stellt das Superoxid-Radikal-Anion, O2–, eine Verbindung
mit einem ungepaarten Valenzelektron dar. Es entsteht formal aus O2 durch die
Aufnahme eines einzelnen Elektrons (univalente Reduktion). Bei saurem pH
assoziert das Superoxid-Radikal-Anion mit einem Proton zum ebenfalls
radikalischen Superoxid. Die Peroxide repräsentieren die nächste Reduktionsstufe
des Sauerstoffs. Obwohl Peroxide keine ungepaarten Valenzelektronen besitzen, sind
sie immer noch erheblich reaktionsfreudig.
Wasserstoffperoxid, H2O2, kann direkt als Nebenprodukt enzymatischer Reaktionen
z.B. der Monoaminoxidase und der Uratoxidase oder indirekt durch Dismutation von
O2– nach folgender Gleichung entstehen:
2 .O2– + 2H+ —› O2 + H2O2
Diese spontane Dismutation läuft bei saurem pH beschleunigt ab.
Ein weiteres Spektrum unterschiedlichster Hydroperoxide kann durch Autooxidation
der ungesättigten Fettsäuren in Membranlipiden entstehen. Solche
Autooxidationsvorgänge finden bei jeglicher Belastung des Organismus mit
Oxidantien, insbesondere mit oxidierenden Radikalen statt. Verschiedene organische
Hydroperoxide werden aber auch in spezifischen enzymatischen Reaktionen
synthetisiert wie z.B die Primärprodukte der Cyclooxygenase- und Lipooxygenase-
Reaktion, PGG2 bzw. 5-HPETE und 15-HPETE.
Das Superoxid-Radikal-Anion entsteht als Nebenprodukt der mitochondrialen
Atmung, als Nebenprodukt von Mikrosomen und wird von zahlreichen Flavin-
abhängigen Dehydrogenasen gebildet. Pathophysiologisch relevant ist die Bildung
Page 7
7
von O2– durch die Xanthinoxidase, die im Gewebe überwiegend als
Xanthindehydrogenase vorliegt, in postischämischem Gewebe.
Xanthindehydrogenase wird unter Ischämiebedingungen durch eine Ca++-abhängige
Protease in Xanthinoxidase überführt. Gleichzeitig kumuliert durch ATP-
Deamidierung das Substrat der Xanthinoxidase, Hypoxanthin. Bei Wiederangebot
von Sauerstoff produziert die zur Xanthinoxidase entartete Xanthindehydrogenase
das Superoxid-Radikal-Anion (Lacy et al., 1998).
Dies wird als wesentliche Noxe bei der postischämischen Gewebsschädigung
betrachtet. Allerdings ist der Xanthindehydrogenasegehalt im Gewebe organ- und
speziesabhängig.
Im Rahmen von Entzündungsphänomenen spielen Phagozyten als Quelle von O2– die
entscheidende Rolle.
In Monozyten, Makrophagen und polymorphkernigen Leukozyten wird O²–durch
eine membranständige NADPH-Oxidase gebildet und ins extrazelluläre Milieu
abgegeben.
NADPH + H+ + 2O2 NADPH-Oxidase › NADP+ + 2.O2– + 2H+
Dabei hat sich der Tumorpromotor Phorbolmyristatacetat, PMA, als ein potenter
Stimulator der phagozytären O2– - Bildung erwiesen (Baum et al., 1998).
Die O2– - Bildung durch Phagozyten ist normalerweise gering, steigt aber bei
Aktivierung, z.B. während der Phagozytose um mehr als das Zehnfache an
(„oxidative burst“). Nicht nur opsonierte Bakterien, sondern eine Vielzahl von nicht-
infektiösen Fremdkörpern und endogenen oder exogenen Substanzen können in
Phagozyten die O2– - Bildung anregen wie z.B der Komplementfaktor C5a,
Leukotrien B4 und bakterielle leukotaktische Peptide.
Daraus resultiert eine lebenswichtige Rolle der Sauerstoffradikale, nämlich die der
Infektabwehr.
Allerdings ist ihre Überproduktion generell toxisch und übersteigt die Kapazität der
zellulären antioxidativen Abwehrmechanismen, die aerobe Zellsysteme als Schutz
gegen die reaktiven Sauerstoffradikale entwickelt haben.
Zur Inaktivierung reaktiver Sauerstoffspezies stehen dem Organismus u.a.
Superoxid-Dismutasen
Page 8
8
2.O2– + 2H+ SOD › H2O2 + O2
und Katalasen
2H2O2 Katalase › O2 + 2H2O
zur Verfügung.
Weiterhin unterdrücken Tryptophan und Ascorbinsäure die Hydroxylradikal-
bildung, während Histidin die Bildung von Singulett-Sauerstoff vermindert.
Weitere Scavenger, die u.a. auch in dieser Studie Verwendung finden, sind Ebselen,
2-Phenyl-1,2-benzisoelenazol-3(2H)-one, Trolox, 6-Hydroxy-2,5,7,8-tetramethyl-
chroman-2-carboxylsäure, Carotinoide, Katecholamine, einige Antiproteasen und
einige Metalle, wie Mangan, Zink und Acetylcystein (Knudsen et al., 1996).
Sauerstoffradikale können bei Überproduktion und bei Überschreitung der zellulären
antioxidativen Abwehrmechanismen Lipide, Proteine, Enzyme, Polysaccharide und
Nukleinsäuren schädigen. Bekanntermaßen schädigen sie Zellmembranen durch die
Peroxidation von ungesättigten Fettsäuren (Halliwell B, 1987).
Desweiteren spielen Sauerstoffradikale eine Rolle bei vielen Erkrankungen, wie z.B.
dem Diabetes mellitus, chronisch inflammatorischen Erkrankungen, dem
Alterungsprozeß, neurologischen Erkrankungen, genetischen Mutationen, der
Atherosklerose, der rheumatoiden Arthritis, neoplastischen Transformationen, der
retrolentalen Fibroplasie bei Neugeborenen und bei der Entstehung des „adult
respiratory distress syndrome“, ARDS (Canavese et al., 1997).
Nicht zuletzt wird Sauerstoffradikalen eine Rolle bei toxischen Nephropathien,
akutem Nierenversagen und der Progression einer Niereninsuffizienz zugeschrieben
(Yoshioka et al., 1990).
Dialysepatienten leiden ebenfalls an einer erhöhten Produktion von reaktiven
Sauerstoffradikalen (Fiorillo et al., 1998).
In den ersten Minuten der Dialyse werden komplementgesteuert polymorphe
mononukleäre Leukozyten an bestimmten Dialysemembranen aktiviert, die zu einer
erhöhten Produktion von reaktiven Sauerstoffradikalen führen (Luciak et al., 1991,
Schettler et al., 1998).
Durch die Aktivierung von neutrophilen, eosinophilen Granulozyten, Monozyten und
Makrophagen kommt es zu einem „respiratory burst“.
Page 9
9
Damit sind Dialysepatienten u.a. dem erhöhten Risiko ARDS-ähnlicher Symptome
ausgesetzt (Hultqvist et al., 1997).
Dialysepatienten leiden durch häufige Blutentnahmen, Punktionen und durch
Retention von Blut im Dialysator unter einem ständigen Blutverlust. So werden bei
jeder Dialyse 15 bis 25ml Blut im Dialysator retiniert (Faich und Strobos, 1999). In
der Vergangenheit wurde die renale Anämie noch durch häufige Bluttransfusionen
behandelt. Durch die häufigen Bluttransfusionen waren die Patienten einem erhöhten
Übertragungsrisiko von Infektionen und einem erhöhten Risiko von Blutgruppen-
inkompatibilitäten ausgesetzt. Heute ist an deren Stelle vorwiegend der Einsatz von
rekombinantem, humanem Erythropoetin getreten. Mit dem Einsatz von EPO und
einem jährlichen Verlust von 6 bis 8g Eisen bei Dialysepatienten ist somit der Bedarf
an Eisen gestiegen (Faich und Strobos, 1999). Erythropoetin stimuliert die Erythro-
poese und damit wird Eisen vermehrt vom Knochenmark aufgenommen, da es für
die Hämoglobinsynthese benötigt wird. Eine orale Eisentherapie hat sich dabei als
uneffektiv erwiesen. So erfolgt die Eisensubstitution bei Dialysepatienten durch die
intravenöse Gabe einer Ampulle Eisen-III-natrium-gluconat-Komplex (Ferrlecit) zu
5 ml oder zu 3,2ml.
Eine Ampulle zu 5ml enthält 62,5mg Eisen-III-natrium-gluconat-Komplex und 45mg
Benzylalkohol, während eine Ampulle zu 3,2ml 28,8mg Benzylalkohol und 40mg
Eisen-III-natrium-gluconat-Komplex enthält.
Dialysepatienten erhalten 3,2ml bzw. 5ml Eisen-III-natrium-gluconat-Komplex je
nach Höhe des Eisendefizits langsam intravenös injiziert oder mit 100ml bis 250ml
physiologischer Kochsalzlösung verdünnt über 20 bis 30 Minuten intravenös
infundiert.
Eisen-III-natrium-gluconat-Komplex, das ein Molekulargewicht von 350.000 Dalton
aufweist, hat sich bei der Behandlung von Dialysepatienten in Bezug auf die
Normalisierung des Hämoglobins, des Hämatokrits, der Eisensättigung und des
Serumferritins als sehr wirksam erwiesen (Nissen et al., Matzke et al., 1999).
Ferner wird von den gleichnamigen Autoren eine viel niedrigere Allergie-und
Nebenwirkungsrate von Eisen-III-natrium-gluconat-Komplex im Vergleich zu
Eisendextran beschrieben.
Page 10
10
Manche Autoren weisen auf ein erhöhtes kardiovaskuläres Risiko und eine erhöhte
Infektionsrate in Zusammenhang mit einer intravenösen Eisentherapie hin (Burns
und Pomposelli, 1999).
Diese Befürchtungen sind laut anderen Studien (Eschbach und Adamson,1999) nicht
nachhaltbar.
Der Eisenbestand eines gesunden Erwachsenen beträgt 3-5g. Von dieser insgesamt
im Körper vorhandenen Eisenmenge ist die Hauptmenge (3/4) in Hämoglobin
gebunden. Eisen ist ein essentieller Bestandteil des Organismus und ist im
Hämoglobin beim Sauerstofftransport, in den Cytochromen beim
Elektronentransport der Atmungskette und in Enzymen bei Oxidations-und
Reduktionsvorgängen beteiligt. Im Blut liegt Eisen nur an Transferrin gebunden vor
(70-180mg/100ml Plasma), das durch die Bindung des dreiwertigen Eisens an Apo-
Transferrin entsteht.
Nach Bindung an spezifischen Membranrezeptoren wird Transferrin zusammen mit
diesen durch Endozytose in die Zelle aufgenommen.
Die Speicherform des Eisens stellt das Ferritin dar, das aus dreiwertigem Eisen
besteht. Ferritin besteht aus einer Proteinschale (Apoferritin) und einem Kern aus
Ferrihydroxyd-Phosphat-Micellen. Ferritin speichert Eisen in biologischer Form und
schützt die Zellen vor der toxischen Wirkung ionisierten Eisens. Das Eisen kann nur
über die Spaltung des Ferritins in Apoferritin und Eisen wieder verwertet werden.
Das Ferritin-gebundene Eisen muß vor seiner Freisetzung wieder zu zweiwertigem
Eisen reduziert werden.
Freies Eisen, das in der zwei-oder dreiwertigen Form vorkommt, ist zwar für viele
biochemische Prozesse lebenswichtig, gilt aber als Katalysator der Fenton (1) - und
der Haber-Weiss-Reaktion (2,3) und wird damit für die Bildung von
Hydroxylradikalen verantwortlich gemacht (Baliga et al., 1997).
1) Fe²+ + H2O2 —› Fe³+ + OH· + OH–
2) Fe³+ + O2– —› Fe²+ + O2
3) Fe²+ + H2O2 —› Fe³+ + OH· + OH–
Auch wird Eisen eine Rolle bei der Progression eines chronischen Nierenversagens,
das mit einer erhöhten Sauerstoffradikalbildung einhergeht, zugeschrieben (Alfrey,
1994).
Page 11
11
Allerdings wurde die Wirkung von Eisen, insbesondere von Eisen-III-natrium-
gluconat-Komplex, auf die Sauerstoffradikalbildung bei Dialysepatienten bisher
nicht gemessen.
Page 12
12
2. PROBLEMSTELLUNG
Die Gabe von intravenösem Eisen-III-natrium-gluconat-Komplex bei Patienten mit
terminaler Niereninsuffizienz, die eine Dialysetherapie erhalten, ist notwendig, um
den erhöhten Eisenbedarf bei gleichzeitiger Gabe von rekombinantem, humanem
Erythropoetin zu decken (Bailie et al., 2000).
Eisen-III-natrium-gluconat-Komplex enthält dreiwertiges Eisen, das als Katalysator
bei der Bildung von Hydroxylradikalen, nach der Haber-Weiss-Reaktion gilt
(Baliga et al., 1997). Mehrfach wurde in Studien zuvor die gesteigerte Bildung von
Sauerstoffradikalen („Oxidative stress“) bei komplexen Prozessen des akuten oder
chronischen Nierenversagens sowie durch die Dialyse selbst beschrieben (Canavese
et al., 1987).
Die Sauerstoffradikalbildung von mononukleären Leukozyten aus dem Blut von
Patienten mit terminaler Niereninsuffizienz läßt sich mit Hilfe eines
Fluoreszenzspektrophotometers messen.
Für die vorliegende Arbeit zur Untersuchung der Wirkung von Eisen-III-natrium-
gluconat-Komplex auf die Sauerstoffradikalbildung von Dialysepatienten ergeben
sich dabei folgende Punkte:
1. Messung der Sauerstoffradikalbildung in vitro bei Dialysepatienten durch Zugabe
von Oxidantien, Antioxidantien und Eisen-III-natrium-gluconat-Komplex
2. Wiederholung dieser Messung bei denselben Patienten nach Gabe von Eisen-III-
natrium-gluconat-Komplex in einem Abstand von zwei Wochen
3. Erstellung einer Dosiswirkungskurve für Eisen-III-natrium-gluconat-Komplex
bei Dialysepatienten und als Vergleich bei gesunden Probanden
4. Vergleich der Wirkung von Fe²+ und Fe³+ auf die Sauerstoffradikalbildung
5. Klärung des Mechanismus bezüglich der Wirkung von Eisen-III-natrium-
gluconat-Komplex
Page 13
13
3. MATERIALIEN
3.1 MATERIALIEN UND REAGENZIEN
Von der Firma Rhône-Poulenc Rorer:
Eisen-III-natrium-gluconat-Komplex (Ferrlecit)
Von der Firma Sigma Chemical Co. (St. Louis):
Superoxiddismutase (SOD), Katalase, 2-Phenyl-1,2-benzisoselenazol-3(2H)-one;
PZ51 (Ebselen), Tyrphostin A51, Captopril, Nigericin, dreiwertiges Eisenchlorid
(FeCl3), zweiwertiges Eisenchlorid (FeCl2), Digitonin, Pluronic F-127, Histopaque-
1077
Von der Firma Merck (Darmstadt):
Natriumazid ( NaN3 ), EXP 3174 (Losartan)
Von der Firma Calbiochem (La Jolla, CA):
Phorbol-12-myristat-13-acetat (PMA), 2´,7´-Dichlorofluoreszindiacetat (DCFA),
Vanadate, Disulfiram, 1,2 Dioctanoyl-sn-glycerol (DOG), 6-Hydroxy-2,5,7,8-
tetramethylchroman-2-carboxylsäure (Trolox), Phen green
Präzisions-Küvetten aus Quarzglas Suprasil von Hellma;
Zentrifugenröhrchen von neoLab Migge Laborbedarf-Vertriebs GmbH, Heidelberg;
Transferpipetten von Sarstedt, Nürnbrecht;
Reaktionsgefäße und Serologische Pipetten, 10 ml, von Carl Roth GmbH und co.,
Karlsruhe;
Microliterspritzen von Hamilton, Schweiz;
Zentrifugenbehälter, 50 ml, San Rafael, USA
Page 14
14
3.2 GERÄTE
Zentrifuge, Laborfuge II, Heraeus Christ (1000-6000 U/min), Osterode/Harz,
Deutschland;
Fluoreszenzspektrophotometer, Modell F-2000 von Hitachi, Tokyo, Japan
pH-Meter, Wissenschaftliche Werkstätten, Weilheim, Deutschland
3.3 LÖSUNGEN
„Hank's balanced salt solution“ (HBSS)
136,00 mM NaCl2 17,94 g/l
5,40 mM KCl 0,40 g/l
0,44 mM KH2PO4 0,06 g/l
0,34 mM Na2HPO4 × 2 H20 0,06 g/l
5,60 mM D-Glucose 1,00 g/l
1,00 mM CaCl2 0,147 g/l
1,00 mM MgCl2 0,203 g/l
10,00 mM HEPES 2,383 g/l
ad 1000 ml Aqua dest.
Die Einstellung erfolgte mit 1 M Natronlauge auf pH 7,4.
Page 15
15
4. METHODEN
4.1 PATIENTENCHARAKTERISIERUNG
Zur Messung von reaktiven Sauerstoffradikalen in mononukleären Leukozyten
dienten neunzehn Probanden als Spender von mononukleären Leukozyten.
Bei sechzehn der Probanden handelte es sich um Dialysepatienten mit terminaler
Niereninsuffizienz, die eine reguläre Dialysebehandlung erhielten.
Die Blutentnahme erfolgte aus dem arteriellen Schenkel des Dialysekreislaufs in eine
20ml umfassende Einmalspritze. Zur Gerinnungshemmung wurde Heparin zugesetzt.
Neben Eisen-III-gluconat erhielten die Dialysepatienten 3*2000 IE Erythropoetin.
Neun Dialysepatienten wurde zweimal 20ml Blut vor und nach Gabe von Eisen-III-
gluconat in Abständen von zwei Wochen vor der Dialyse entnommen.
Bei den restlichen sieben Dialysepatienten erfolgte die Blutentnahme einmalig.
Die Patienten, davon neun weiblich und sieben männlich, waren zwischen fünfzig
und neunzig Jahre alt (Mittelwert mit Standardabweichung: 65,44±4,7).
Die Ursachen der terminalen Niereninsuffizienz waren eine diabetische
Nephropathie bei sechs Patienten, eine Nephrosklerose bei vier Patienten, eine
interstitielle Nephritis bei drei und eine Glomerulonephritis bei zwei Patienten.
Bei vier Patienten konnte die Ursache der terminalen Niereninsuffizienz nicht eruiert
werden.
Zwölf der Dialysepatienten wiesen eine koronare Herzkrankheit (KHK), zwei eine
obstruktive Lungenkrankheit und sechs eine periphere arterielle Verschlußkrankheit
(pAVK) auf.
Drei Spender von mononukleären Leukozyten waren gesunde Kontrollpersonen, bei
denen keine Erkrankung bekannt war. Sie waren zwischen zwanzig und vierzig Jahre
alt (Mittelwert und Standardabweichung: 28,34±5,93).
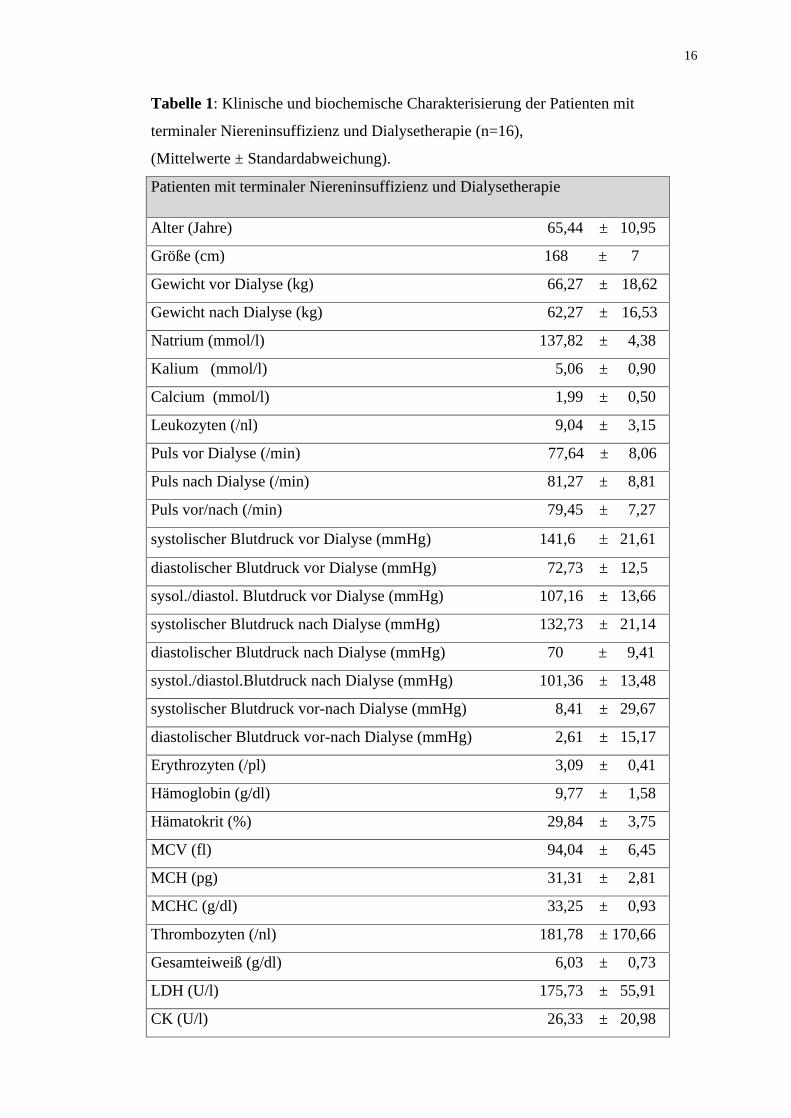
Die klinische und biochemische Charakterisierung der Patienten ist in der Tabelle 1
aufgeführt.
Page 16
16
Tabelle 1: Klinische und biochemische Charakterisierung der Patienten mit
terminaler Niereninsuffizienz und Dialysetherapie (n=16),
(Mittelwerte ± Standardabweichung).
Patienten mit terminaler Niereninsuffizienz und Dialysetherapie
Alter (Jahre) 65,44 ± 10,95
Größe (cm) 168 ± 7
Gewicht vor Dialyse (kg) 66,27 ± 18,62
Gewicht nach Dialyse (kg) 62,27 ± 16,53
Natrium (mmol/l) 137,82 ± 4,38
Kalium (mmol/l) 5,06 ± 0,90
Calcium (mmol/l) 1,99 ± 0,50
Leukozyten (/nl) 9,04 ± 3,15
Puls vor Dialyse (/min) 77,64 ± 8,06
Puls nach Dialyse (/min) 81,27 ± 8,81
Puls vor/nach (/min) 79,45 ± 7,27
systolischer Blutdruck vor Dialyse (mmHg) 141,6 21,61
diastolischer Blutdruck vor Dialyse (mmHg) 72,73 ± 12,5
sysol./diastol. Blutdruck vor Dialyse (mmHg) 107,16 ± 13,66
systolischer Blutdruck nach Dialyse (mmHg) 132,73 ± 21,14
diastolischer Blutdruck nach Dialyse (mmHg) 70 ± 9,41
systol./diastol.Blutdruck nach Dialyse (mmHg) 101,36 ± 13,48
systolischer Blutdruck vor-nach Dialyse (mmHg) 8,41 ± 29,67
diastolischer Blutdruck vor-nach Dialyse (mmHg) 2,61 ± 15,17
Erythrozyten (/pl) 3,09 ± 0,41
Hämoglobin (g/dl) 9,77 ± 1,58
Hämatokrit (%) 29,84 ± 3,75
MCV (fl) 94,04 ± 6,45
MCH (pg) 31,31 ± 2,81
MCHC (g/dl) 33,25 ± 0,93
Thrombozyten (/nl) 181,78 ± 170,66
Gesamteiweiß (g/dl) 6,03 ± 0,73
LDH (U/l) 175,73 ± 55,91
CK (U/l) 26,33 ± 20,98
Page 17
17
Kalium (mmol/l) 5,15 ± 0,84
Calcium (mmol/l) 1,97 ± 0,52
Phosphate (mg/dl) 4,82 ± 2,24
Chlorid (mmol/l) 103 ± 0
Glucose (mg/dl) 141,8 ± 17,61
Ultrafiltrationsrate (ml) 1273,57 ± 1340,47
Kreatinin (mg/dl) 6,24 ± 2,34
Harnstoff (mg/dl) 134,63 ± 53,32
Harnsäure (mg/dl) 4,9 ± 1,81
?GT (U/l) 37,29 ± 30,16
GPT (U/l) 13,7 ± 5,05
GOT(U/l) 13,45 ± 3,87
Cholesterin (mg/dl) 186,56 ± 44,45
Triglyceride (mg/dl) 311,5 ± 245,38
Dialysedauer (/h) 4,08 ± 0,74
Leitwert (ms/cm) 14,13 ± 0,2
Page 18
18
4.2 PRINZIP DER FLUORESZENZ-SPEKTROPHOTOMETRIE
Die Fluoreszenzmessung besteht darin, spezifische Farbstoffmoleküle in einen
angeregten Zustand zu überführen. Die Anregung erfolgt durch Energiezufuhr mit
Hilfe einer Xenon-Lampe. Die Elektronen der Farbstoffmoleküle werden auf
energiereichere Bahnen angehoben. Während die Moleküle aus dem angeregten
Zustand in den Grundzustand zurückkehren, wird die freiwerdende Energie in Form
von elektromagnetischen Lichtquanten definierter Wellenlängen emittiert.
Dies bezeichnet man als Fluoreszenz (Haeckel & Schumann). Die Xenon-Lampe
setzt eine Primärstrahlung (Exzitationsstrahlung) aus, die durch einen Filter (Ex-
zitationsfilter) verändert wird. Dieser Filter läßt nur eine bestimmte Wellenlänge
hindurch. Trifft die Exzitationsstrahlung auf die Lymphozytenprobe, werden die
Moleküle in einen höheren Energiezustand versetzt, fallen wieder in den
Grundzustand zurück und setzen dabei Strahlung frei. Bevor diese Strahlung auf
einen Detektor trifft, der diese in eine Meßzahl umwandelt, passiert sie einen
Emissionsfilter, der nur eine bestimmte Strahlung durchläßt.
Die Intensität der mit Hilfe des Fluoreszenz-Spektrophotometers gemessenen
Fluoreszenz ist dabei von der Anzahl der in einer Probe sich befindlichen Moleküle
abhängig.
4.3 PRÄPARATION VON MONONUKLEÄREN LEUKOZYTEN
Die mononukleären Leukozyten wurden aus heparinisierten Blutröhrchen in
folgender Art und Weise gewonnen (Nelson et al.,1999):
Zuerst wurden die Blutröhrchen mit 150 * g für 15 Minuten zur Trennung des
Blutplasmas zentrifugiert, das Plasma wurde verworfen und der 5ml enthaltende
Zellbestand mit „Hank´s balanced salt solution“, HBSS, 1:1 verdünnt. Dieser
Zellbestand wurde nun in jeweils 3ml enthaltende Histopaque-Röhrchen pipettiert
und mit
230 * g für 15 Minuten zentrifugiert. Die mittlere Phase der mononukleären
Leukozyten wurde sorgfältig aspiriert, mit HBSS auf 10ml aufgefüllt und mit 230 * g
für 5 Minuten erneut zentrifugiert.
Page 19
19
Der Überstand wurde verworfen und der Rest mit 3ml HBSS, und 50µl des
Farbstoffes Dichlorofluoreszindiacetat, DCFDA, für 10 Minuten in abge-
dunkelter Umgebung inkubiert.
Nach erneuter Zentrifugation mit 230 * g für 5 Minuten wurden die mononukleären
Leukozyten mit 10ml HBSS verdünnt. Davon wurden 1000µl in 10 Küvetten
meßfertig gefüllt.
4.4 MESSUNG VON MONONUKLEÄREN LEUKOZYTEN
Dichlorofluoreszindiacetat, DCFDA, ist eine apolare Verbindung, die leicht in Zellen
diffundiert, wo es in die nicht fluoreszierende polare Form DCFH hydrolisiert wird.
Damit bleibt es als letztere Form in den Zellen gefangen. In Anwesenheit von
Sauerstoffradikalen, ROS, wird DCFH in die stark fluoreszierende Form 2',7'-
dichlorofluoreszin (DCF) oxidiert (Nelson et al., 1999).
Die Fluoreszenz von DCF wird durch Radikale wie das Superoxidanion, das
Hydroxyl-Ion, Hydrogenperoxid und durch den Singulett-Sauerstoff gesteigert
(Watanbe, 1998). Die Höhe der Fluoreszenz von DCF, die die Konzentration von
vorhandenen Sauerstoffradikalen wiederspiegelt (Le Bel et al., 1992), wurde
spektrophotometrisch bei einer Wellenlänge von 534nm und 488nm mit dem
Spektrophotometer F-2000 Hitachi, Tokyo, Japan, gemessen.
Da die Fluoreszenz nur eine Minute lang in bestimmten Zeitintervallen von 0, 30 und
60 Minuten gemessen wurde, konnten die Effekte der Photooxidation von DCFH
minimiert werden (Nelson et al., 1999).
PMA, Phorbol-12-myristat-13-acetat, ein Aktivator der Proteinkinase C (Rehan et
al., 1985), wurde in einer Konzentration von 100nmol/L benutzt, um die sich in
Küvetten befindlichen mononukleären Leukozyten zu aktivieren.
4.5 MESSUNG MIT OXIDANTIEN UND ANTIOXIDANTIEN
Die Messung von Sauerstoffradikalen nach 0, 30 und 60 Minuten erfolgte in der
Anwesenheit von Superoxiddismutase (40µg/mL), einem Metallprotein, das
Page 20
20
Superoxid-Radikale in H2O2 umwandelt (McCord, 1986), Catalase (5µg/mL), einem
Hämprotein (Escobar et al., 1996), das H2O2 zu H2O und O2 umbaut, Ebselen, 2-
Phenyl-1,2-benzisoselenazol-3(2H)-one (10 µmol/L), einem Peroxy-nitritfänger
(Husain et al., 1997), NaN3, Natriumazid (1mmol/L), einem Singulettsauer-
stofffänger, Tyrphostin A51 (10µmol/L), einem Tyrosinkinase-inhibitor (Novo-
grodsky et al., 1998), Captopril (100µmol/L), einem Hemmer des Angiotensin-
Converting-Enzyms (Aruoma et al., 1991), EXP 3174, einem aktiven Metaboliten
des Losartan (100µg/µL), einem Angiotensin-II-RezeptorAntagonisten (Tamaki et
al., 1993) und Eisen-III-natrium-gluconat-Komplex (31,25µg/mL).
Bei jeder Messung wurde die Fluoreszenz von mononukleären Leukozyten ohne die
Anwesenheit von PMA, Phorbol-12-myristat-13-acetat, als Kontrollwert gemessen.
Zur Erstellung einer Dosiswirkungskurve für Eisen-III-natrium-gluconat-Komplex
wurde die Fluoreszenz des Farbstoffes Dichlorofluoreszindiazetat, dessen Höhe den
Gehalt an Sauerstoffradikalen anzeigt (Le Bel et al., 1992), ebenfalls nach 0, 30 und
60 Minuten, in Anwesenheit von PMA allein und in Anwesenheit von Eisen-III-
natrium-gluconat-Komplex bestimmt. Eisen-III-natrium-gluconat-Komplex wurde
dabei unterschiedlich dosiert. Es wurden jeweils 1µl, 5µl, 10µl, 20µl, 50µl und 100µl
Eisen-III-natrium-gluconat-Komplex in sechs Küvetten zusammen mit dem
Stimulator PMA (10µl) pipettiert.
Aus den Meßzahlen von PMA allein und PMA zusammen mit Eisen-III-natrium-
gluconat-Komplex in sechsmal unterschiedlicher Dosierung ergab sich nun eine
Dosiswirkungskurve.
Diese wurde mit Hilfe von gewonnenen mononukleären Leukozyten von drei
Dialysepatienten und zum Vergleich mit Hilfe von mononukleären Leukozyten von
drei gesunden Probanden erstellt.
Desweiteren erfolgte die Erstellung einer Zeitverlaufskurve über 30 Minuten bei
488/534nm getrennt für PMA und für PMA zusammen mit 50µl Eisen-III-natrium-
gluconat-Komplex.
Bei diesem Versuch wurden präparierte mononukleäre Leukozyten von
Dialysepatienten verwendet.
Page 21
21
4.6 MESSUNG MIT VANADATE, DISULFIRAM, DOG UND EISEN-III-
NATRIUM-GLUCONAT-KOMPLEX
Die Messung der Fluoreszenz erfolgte mit 10µl PMA allein, 10µl PMA und 50µl
Eisen-III-natrium-gluconat-Komplex, 10µl PMA, 50µl Eisen-III-natrium-gluconat-
Komplex und 10µl Vanadate, 10µl PMA und 10µl Vanadate, 10µl PMA, 50µl Eisen-
III-natrium-gluconat-Komplex und 20µl Disulfiram, 10µl DOG, 1,2-Dioctanoyl-sn-
glycerol, allein und 10µl DOG mit 50µl Eisen-III-natrium-gluconat-Komplex.
Vanadate ist ein Hemmer von ATPasen, Phosphotransferasen- und kinasen
(Monteiro et al., 1991). Ferner stimuliert es die NADP-Oxidase und begünstigt damit
die Sauerstoffradikalbildung (Carmichael A, 1995).
DOG ist ein Aktivator der Proteinkinase C und ist der Wirkung des Tumorpromotors
PMA, der Proteinkinase C, sehr ähnlich (Baum et al., 1998).
Disulfiram stellt einen Inhibitor der Alkohol-und Aldehyddehydrogenase I +II und
einen Inhibitor der Caspase-1 und Caspase-3 dar.
Disulfiram senkt die Sauerstoffradikalbildung (Koksoy et al., 1997).
4.7 MESSUNG MIT FeCl3, FeCl2 UND TROLOX
Zur Mechanismusaufklärung wurden präparierten mononukleären Leukozyten aus
Patientenblut, bei denen eine Dialysetherapie erfolgte, 10µl PMA allein, 10µl PMA
mit 50µl FeCl3, 10µl PMA mit 50µl FeCl2 und 10µl PMA mit 10µl Trolox zugesetzt.
Anschließend wurde auch hier die Fluoreszenz und damit die Sauerstoffradikal-
bildung gemessen.
Trolox, 6-Hydroxy-2,5,7,8-tetramethylchroman-2-carboxylsäure ist ein wasser-
lösliches Vitamin E-Derivat mit sehr potenten antioxidativen Eigenschaften
(Evstigneva et al., 1998).
FeCl3 und FeCl2 enthalten zwei-und dreiwertiges Eisen und gelten als Katalysatoren
der Fenton/Haber-Weiss-Reaktion, damit der Sauerstoffradikalbildung und daher als
potentiell schädlich (Baliga et al., 1997).
Page 22
22
4.8 VERSUCHE MIT DEM EISENINDIKATOR PHEN GREEN
Phen green ist in der Lage zweiwertiges Eisen, FeCl2, zu binden und damit die
Anwesenheit von zweiwertigem Eisen zu messen (Petrat et al., 2000).
Die Intensität der durch phen green gemessenen Fluoreszenz ist dabei sowohl von
der Konzentration des Indikators selbst als auch von der Konzentration von
zweiwertigem Eisen abhängig.
Nach der Präparation von mononukleären Leukozyten wurden diese mit 10µl der
Stammlösung von phen green für 20 Minuten in abgedunkelter Umgebung inkubiert.
Anschließend wurden die mononukleären Leukozyten 5 Minuten lang bei 230*g
zentrifugiert. Nach der Zentrifugation wurde der Überstand verworfen und die
mononukleären Leukozyten wurden mit 6 ml HBSS aufgefüllt.
Danach wurden jeweils 1000µl der Lösung von mononukleären Leukozyten in 6
Küvetten meßfertig gefüllt. Die Endkonzentration von phen green in den Küvetten
betrug 10µmol/l.
Die Messung der Fluoreszenz in jeder Küvette betrug 400 Sekunden. Nach 200
Sekunden wurden den mit phen green inkubierten mononukleären Leukozyten in der
ersten Küvette 10µl Digitonin zugesetzt. Die Endkonzentration von Digitonin in den
Küvetten betrug 20mmol/l.
Der zweiten Küvette wurden nach 200 Sekunden 100µl FeCl2 zugesetzt. Die
Endkonzentration von FeCl2 in der Küvette betrug 10mol/l.
In der dritten Küvette wurden den mononukleären Leukozyten, die vorher zusätzlich
mit 50µl Eisen-III-natrium-gluconat-Komplex für 30 Minuten inkubiert wurden,
nach 200 Sekunden erneut 10µl Digitonin zugesetzt. Der vierten Küvette wurde der
Lösung von mononukleären Leukozyten, die vorher mit 50µl Eisen-III-natrium-
gluconat-Komplex für 30 Minuten inkubiert wurde, 100µl FeCl2 zugesetzt.
Auch der fünften und sechsten Küvette wurde den mononukleären Leukozyten nach
200 Sekunden zunächst Digitonin, dann FeCl2 zugesetzt.
Die fünfte und sechste Küvette wurden vorher für 60 Minuten mit 50µl Eisen-III-
natrium-gluconat-Komplex inkubiert.
Nach jeder Messung erhielt man eine Kurve, an der zuerst der Basalwert und dann
nach Zugabe von Digitonin oder FeCl2 die veränderte Fluoreszenz abgelesen wurde.
Jeder Schritt wurde jeweils dreimal wiederholt.
Page 23
23
4.9 KONTROLLVERSUCHE IN ABWESENHEIT VON
MONONUKLEÄREN LEUKOZYTEN
Eisen-III-natrium-gluconat-Komplex konnte in den zuvor beschriebenen Versuchen
in Anwesenheit von mononukleären Leukozyten die PMA-induzierte
Sauerstoffradikalbildung hemmen, sofern die mononukleären Leukozyten von
Dialysepatienten stammten. Die Hemmung der Sauerstoffradikalbildung durch
Eisen-III-natrium-gluconat-Komplex resultiert dabei aus einer Wirkung auf die
mononukleären Leukozyten selbst. Um zu zeigen, daß die Verminderung der
Fluoreszenz durch Eisen-III-natrium-gluconat-Komplex nicht etwa auf einer
Wirkung auf den Farbstoff Dichlorofluoreszindiacetat beruht, wurden Versuche in
Abwesenheit von mononukleären Leukozyten durchgeführt.
Zunächst wurde die Fluoreszenz von 1000µl „Hank´s balanced salt solution“, HBSS,
allein, dann mit 1000µl HBSS und 10µl Eisen-III-natrium-gluconat-Komplex und
anschließend mit 1000µl HBSS, 10µl Eisen-III-natrium-gluconat-Komplex und
1000µl HBSS gemessen.
Weiterhin erfolgte die Messung der Fluoreszenz mit 1000µl HBSS allein, 1000µl
HBSS und 10µl Eisen-III-natrium-gluconat-Komplex und anschließend mit 1000µl
HBSS, 10µl Eisen-III-natrium-gluconat-Komplex und 1000µl H2O2.
Diese Versuchsschritte wurden jeweils fünfmal wiederholt.
Durch Zugabe von HBSS oder H2O2 erfolgte jedoch keine Veränderung der
Fluoreszenz von Eisen-III-natrium-gluconat-Komplex.
Desweiteren erfolgte die Messung des pH-Wertes und der Fluoreszenz von 1000µl
H2O2 allein , dann jeweils unter Zugabe von 10µl DCFDA, von 10µl Eisen-III-
natrium-gluconat-Komplex, von 40µl Eisen-III-natrium-gluconat-Komplex und unter
Zugabe von 50µl Eisen-III-natrium-gluconat-Komplex. Dieser Schritt wurde
zweimal durchgeführt.
Der pH-Wert und die Fluoreszenz wurden ebenfalls von 1000µl HBSS allein und
unter Zugabe von jeweils 10µl DCFDA, 10µl Eisen-III-natrium-gluconat-Komplex,
40µl Eisen-III-natrium-gluconat-Komplex und 50µl Eisen-III-natrium-gluconat-
Komplex gemessen.
Auch dieser Versuchsschritt wurde zweimal durchgeführt.
Page 24
24
Die Fluoreszenz stieg unter Zugabe von DCFDA, blieb aber unter weiterer Zugabe
von Eisen-III-natrium-gluconat-Komplex nahezu unverändert. Der pH-Wert sank
von 7,32 auf 7,12 nach Zugabe des Farbstoffes DCFDA, blieb aber unter weiterer
Zugabe von Eisen-III-natrium-gluconat-Komplex konstant bei 7,14.
Damit konnte gezeigt werden, daß Eisen-III-natrium-gluconat-Komplex keinen
Einfluß auf den Farbstoff DCFDA und ebenfalls keinen Einfluß auf den pH-Wert
hatte.
4.10 STATISTIK
Der U-Test von Wilcoxon, Mann und Whitney prüft, ob sich die Mittelwerte von
zwei unabhängigen Stichproben signifikant oder nur zufällig voneinander
unterscheiden.
Die Nullhypothese lautet:
Ho: µ1 = µ2 = µ
Dieser Test für unverbundene Stichproben ist verteilungsunabhängig. Der Umfang
der einen Stichprobe sei n1 und der anderen Stichprobe sei n2.
Um eine gemeinsame aufsteigende Rangordnung zu erhalten, werden die Daten der
beiden Stichproben zusammengefasst (N = n1+n2) und der Größe nach geordnet. Die
Summe der Rangzahlen von Stichprobe 1 sei R1 und die von Stichprobe 2 sei R2.
Folgende Größen werden als Prüfgrößen verwendet:
U2 = n1 x n2 + ½ x n2 x (n2 + 1) – R2
U1 = n1 x n2 + ½ x n1 x (n1 + 1) – R1
Der kleinere Wert der beiden Prüfgrößen wird als kritischer Wert U betrachtet. Mit
folgender Prüfgleichung erfolgt die Kontrolle, ob richtig gerechnet wurde:
U1 + U2 = n1 x n2
Die Nullhypothese wird mit der Irrtumswahrscheinlichkeit
verworfen, wenn U als
der niedrigere Wert von U1 und U2 kleiner oder gleich dem tabellierten kritischen
Wert Uo (n1, n2, ) ist (Ramm & Hoffman, Biomathematik; Sachs, Angewandte
Statistik).
Als Irrtumswahrscheinlichkeit, für den Fehler 1. Art, wurde < 0,05 festgelegt.
Page 25
25
5. ERGEBNISSE
4.2 WIRKUNG VON OXIDANTIEN UND ANTIOXIDANTIEN
Die Messung von Sauerstoffradikalen nach 0, 30 und 60 Minuten nach Zugabe von
Oxidantien und Antioxidantien erfolgte zwei Wochen vor und zwei Wochen nach
Gabe von Eisen-III-natrium-gluconat-Komplex in vitro in mononukleären
Leukozyten von Patienten mit terminaler Niereninsuffizienz.
Die Messungen erfolgten mit Hilfe des Fluoreszenz-Farbstoffes, Dichlorofluoreszin-
diacetat, DCFDA. Sowohl die basale Sauerstoffradikalbildung als auch die
Produktion von reaktiven Sauerstoffradikalen nach Stimulation der Zellen mit dem
Phorbolester PMA wurden untersucht.
Die basale Sauerstoffradikalbildung war im Mittel 211 ± 51 % des Ausgangs-
wertes (n=9).
Nach Stimulation mit PMA kam es zu einem signifikanten Anstieg der
Sauerstoffradikalbildung von 211 ± 51 % auf 682 ± 145 % des Ausgangswertes
(n=9; p=0.004).
Die basale Sauerstoffradikalbildung nach 14tägiger Gabe von Eisen-III-natrium-
gluconat-Komplex war 231 ± 34 % des Ausganswertes (n=9).
Nach 14tägiger Gabe von Eisen-III-natrium-gluconat-Komplex führte die
Stimulation mit PMA zu einem signifikanten Anstieg der Sauerstoffradikalbildung
von 231 ± 34 % auf 554 ± 194 % des Ausgangswertes (n=9; p=0.049).
Dann wurden die Effekte verschiedener antioxidativ wirksamer Substanzen auf die
Bildung von reaktiven Sauerstoffradikalen in vitro untersucht.
Am stärksten antioxidierend wirkten dabei Ebselen und Natriumazid, NaN3. Die
antioxidative Wirkung von Ebselen wurde schon von vielen Autoren beschrieben
(Husain et al., 1997).
Die Vorgabe von Ebselen führte zu einer signifikanten Verminderung der PMA-
induzierten Produktion von reaktiven Sauerstoffradikalen von 682 ± 145 % auf
229,±32 % (n=9, p=0.002).
In mononukleären Zellen nach 14tägiger Gabe von Eisen-III-natrium-gluconat-
Komplex war die PMA-induzierte Sauerstoffradikalbildung durch die Zugabe von
Ebselen signifikant vermindert von 554±194 % auf 179±60% (n=9, p=0.007).
Page 26
26
Vor Gabe von Eisen-III-natrium-gluconat-Komplex war die PMA-induzierte
Sauerstoffradikalbildung durch die Zugabe von NaN3 signifikant vermindert von
682±145 % auf 299±50% (n=9, p=0.040).
In mononukleären Zellen nach 14tägiger Gabe von Eisen-III-natrium-gluconat-
Komplex war die PMA-induzierte Sauerstoffradikalbildung durch Zugabe von NaN3
ebenfalls signifikant vermindert von 554±194 % auf 242±54 % (n=9, p=0.038).
Auch der Hemmer des Angiotensin-Converting-Enzyms, Captopril, wie auch ein
Angiotensin-II-Rezeptor-Antagonist, EXP 3174, ein aktiver Metabolit des Losartan,
senkten die PMA-induzierte Sauerstoffradikalbildung.
Hemmer des Angiotensin-Converting -Enzyms sowie Angiotensin-II-Rezeptor-
Antagonisten gelten als nephroprotektiv und als Sauerstoffradikalfänger
(Aruoma et al., 1991; Mira et al., 1998; Sweet et al., 1994;Tamaki et al., 1993).
In mononukleären Zellen vor Gabe von Eisen-III-natrium-gluconat-Komplex war die
PMA-induzierte Sauerstoffradikalbildung durch die Vergabe von Captopril
signifikant vermindert und sank von 682±145 % auf 386±77 % (n=9, p=0.004). In
mononukleären Zellen nach 14tägiger Gabe von Eisen-III-natrium-gluconat-
Komplex war die PMA-induzierte Sauerstoffradikalbildung durch die Vergabe von
Captopril ebenfalls signifikant vermindert und sank von 554±194 % auf 301±94 %
(n=9, p=0.055).
In mononukleären Zellen vor Gabe von Eisen-III-natrium-gluconat-Komplex war die
PMA-induzierte Sauerstoffradikalbildung durch die Zugabe von EXP 3174 ebenfalls
vermindert. Die Fluoreszenz konnte von 682±145 % auf 372±112 % (n=9, p=0.027)
gesenkt werden.
In mononukleären Zellen nach 14tägiger Gabe von Eisen-III-natrium-gluconat-
Komplex war die PMA-induzierte Sauerstoffradikalbildung durch Zugabe von EXP
3174 vermindert von 554±194 % auf 156±33 % (n=9, p=0.008).
Die als antioxidierend bekannten Scavenger Catalase und Superoxiddismutase
(Escobar et al., 1996; McCord J, 1986) führten nicht oder nur zu einer geringfügigen
Verminderung der PMA-induzierten Bildung von reaktiven Sauerstoffradikalen.
Vor Gabe von Eisen-III-natrium-gluconat-Komplex führte die Zugabe von
Superoxiddismutase in mononukleären Zellen sogar zu einer Erhöhung der PMA-
Page 27
27
induzierten Produktion von reaktiven Sauerstoffradikalen von 682±145 % auf
803±164 % (n=9, p=0.359).
In mononukleären Zellen nach 14tägiger Gabe von Eisen-III-natrium-gluconat-
Komplex fiel die PMA-induzierte Produktion von Sauerstoffradikalen geringfügig
durch die Zugabe von Superoxiddismutase von 554±194 % auf 472±127 % (n=9,
p=0.547).
In mononukleären Zellen vor Gabe von Eisen-III-natrium-gluconat-Komplex war die
PMA-induzierte Sauerstoffradikalbildung durch die Vergabe von Catalase leicht
erhöht von 682±145% auf 738±131 % (n=9, p=0.496).
In mononukleären Zellen nach 14tägiger Gabe von Eisen-III-natrium-gluconat-
Komplex war die PMA-induzierte Sauerstoffradikalbildung durch die Vergabe von
Catalase ebenfalls leicht erhöht von 554±194 % auf 616±103 % (n=9, p=0.313).
Tyrphostin, ein Tyrosinkinaseinhibitor (Novogrodsky et al., 1998), konnte die
Produktion von reaktiven Sauerstoffradikalen sowohl vor als auch zwei Wochen
nach Gabe von Eisen-III-natrium-gluconat-Komplex in geringem Maße senken.
Die Bildung von PMA-induzierten Sauerstoffradikalen vor Gabe von Eisen-III-
natrium-gluconat-Komplex sank unter Zugabe von Tyrphostin geringfügig von
682±145 % auf 566±126 % (n=9, p=0.250).
In mononukleären Zellen nach 14tägiger Gabe von Eisen-III-natrium-gluconat-
Komplex führte die Zugabe von Tyrphostin zu einer ebenfalls geringfügigen
Verminderung der Bildung von Sauerstoffradikalen von 554±194 % auf 324±124
% (n=9, p=0.008).
Der Eisen-III-natrium-gluconat-Komplex (Ferrlecit) führte in vitro zu einer
Verminderung der PMA-induzierten Sauerstoffradikalbildung.
Die Vorgabe von Eisen-III-natrium-gluconat-Komplex konnte die PMA-induzierte
Sauerstoffradikalproduktion zwei Wochen vor intravenöser Gabe von Eisen-III-
natrium-gluconat-Komplex an die Patienten von 682±145% auf 429±84 % (n=9,
p=0.008) senken.
In mononukleären Zellen nach 14tägiger Gabe von intravenösem Eisen-III-natrium-
gluconat-Komplex führte die Zugabe von Eisen-III-natrium-gluconat-Komplex zu
einer geringfügigen Veränderung der PMA-induzierten Sauerstoffradikalbildung von
554±194 % auf 488±177 % ( n=9, p=0.313 ).
Page 28
28
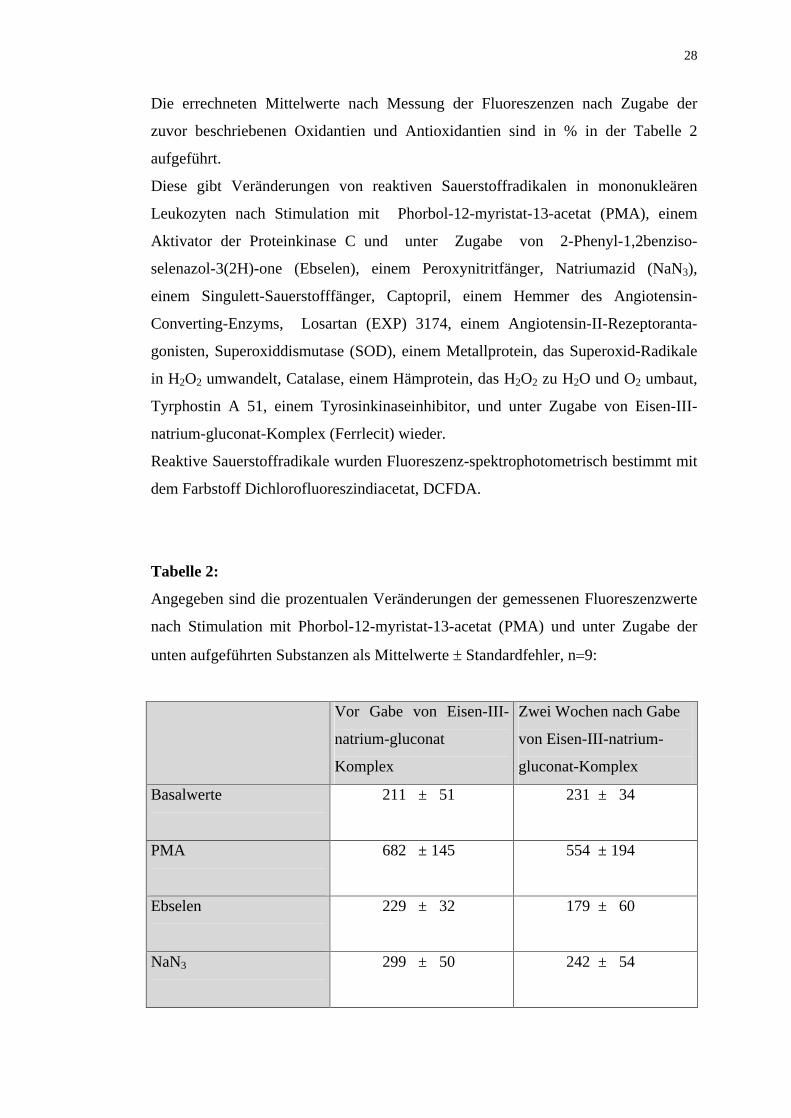
Die errechneten Mittelwerte nach Messung der Fluoreszenzen nach Zugabe der
zuvor beschriebenen Oxidantien und Antioxidantien sind in % in der Tabelle 2
aufgeführt.
Diese gibt Veränderungen von reaktiven Sauerstoffradikalen in mononukleären
Leukozyten nach Stimulation mit Phorbol-12-myristat-13-acetat (PMA), einem
Aktivator der Proteinkinase C und unter Zugabe von 2-Phenyl-1,2benziso-
selenazol-3(2H)-one (Ebselen), einem Peroxynitritfänger, Natriumazid (NaN3),
einem Singulett-Sauerstofffänger, Captopril, einem Hemmer des Angiotensin-
Converting-Enzyms, Losartan (EXP) 3174, einem Angiotensin-II-Rezeptoranta-
gonisten, Superoxiddismutase (SOD), einem Metallprotein, das Superoxid-Radikale
in H2O2 umwandelt, Catalase, einem Hämprotein, das H2O2 zu H2O und O2 umbaut,
Tyrphostin A 51, einem Tyrosinkinaseinhibitor, und unter Zugabe von Eisen-III-
natrium-gluconat-Komplex (Ferrlecit) wieder.
Reaktive Sauerstoffradikale wurden Fluoreszenz-spektrophotometrisch bestimmt mit
dem Farbstoff Dichlorofluoreszindiacetat, DCFDA.
Tabelle 2:
Angegeben sind die prozentualen Veränderungen der gemessenen Fluoreszenzwerte
nach Stimulation mit Phorbol-12-myristat-13-acetat (PMA) und unter Zugabe der
unten aufgeführten Substanzen als Mittelwerte Standardfehler, n 9:
Vor Gabe von Eisen-III-
natrium-gluconat
Komplex
Zwei Wochen nach Gabe
von Eisen-III-natrium-
gluconat-Komplex
Basalwerte
211 ± 51 231 ± 34
PMA
682 ± 145 554 ± 194
Ebselen
229 ± 32 179 ± 60
NaN3
299 ± 50 242 ± 54
Page 29
29
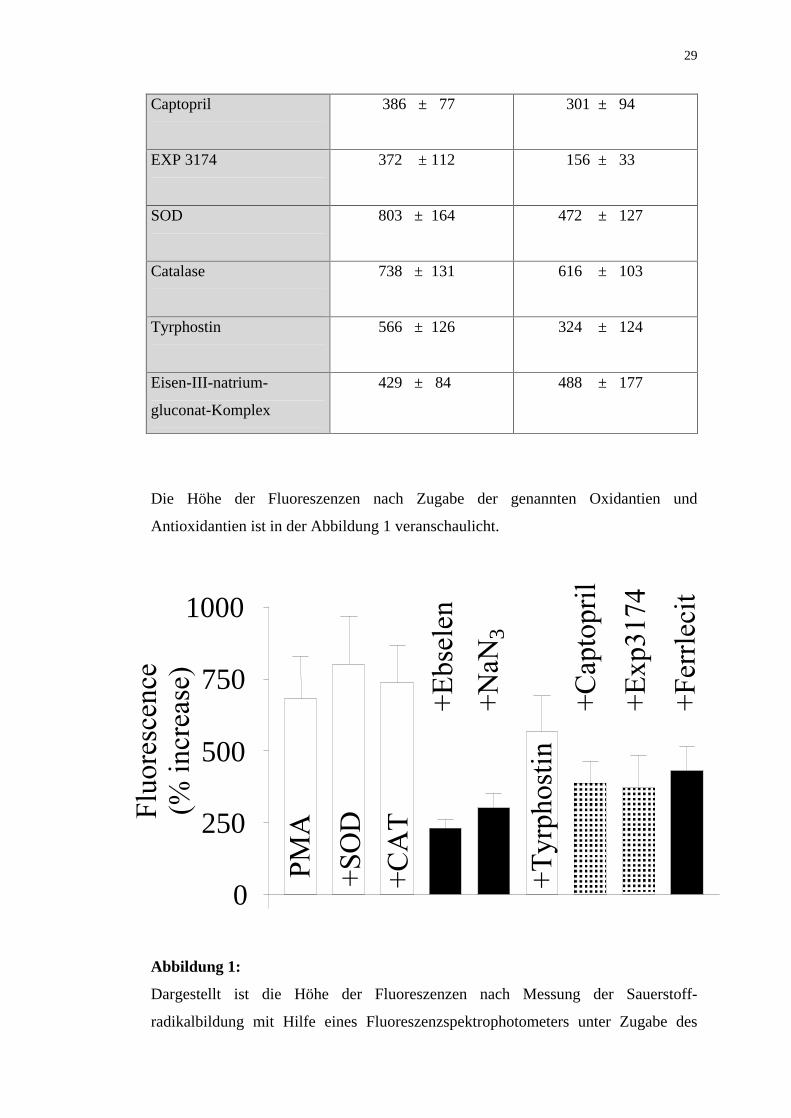
Captopril
386 ± 77 301 ± 94
EXP 3174
372 ± 112 156 ± 33
SOD
803 ± 164 472 ± 127
Catalase
738 ± 131 616 ± 103
Tyrphostin
566 ± 126 324 ± 124
Eisen-III-natrium-
gluconat-Komplex
429 ± 84 488 ± 177
Die Höhe der Fluoreszenzen nach Zugabe der genannten Oxidantien und
Antioxidantien ist in der Abbildung 1 veranschaulicht.
Abbildung 1:
Dargestellt ist die Höhe der Fluoreszenzen nach Messung der Sauerstoff-
radikalbildung mit Hilfe eines Fluoreszenzspektrophotometers unter Zugabe des
0
250
500
750
1000
3
Page 30
30
Farbstoffs 2´,7´-Dichloro-fluoreszindiacetat (DCFDA) und unter der Zugabe der in
der Abbildung aufgeführten Oxidantien und Antioxidantien.
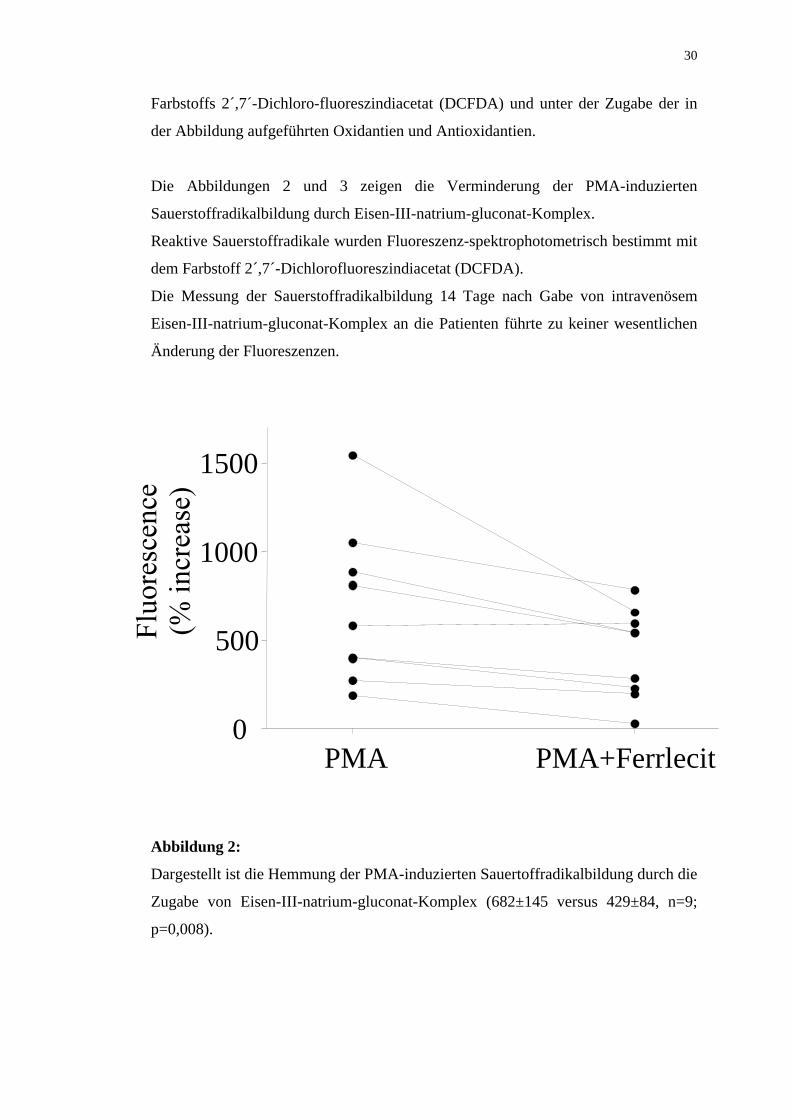
Die Abbildungen 2 und 3 zeigen die Verminderung der PMA-induzierten
Sauerstoffradikalbildung durch Eisen-III-natrium-gluconat-Komplex.
Reaktive Sauerstoffradikale wurden Fluoreszenz-spektrophotometrisch bestimmt mit
dem Farbstoff 2´,7´-Dichlorofluoreszindiacetat (DCFDA).
Die Messung der Sauerstoffradikalbildung 14 Tage nach Gabe von intravenösem
Eisen-III-natrium-gluconat-Komplex an die Patienten führte zu keiner wesentlichen
Änderung der Fluoreszenzen.
Abbildung 2:
Dargestellt ist die Hemmung der PMA-induzierten Sauertoffradikalbildung durch die
Zugabe von Eisen-III-natrium-gluconat-Komplex (682±145 versus 429±84, n=9;
p=0,008).
0
500
1000
1500
PMA PMA+Ferrlecit
Page 31
31
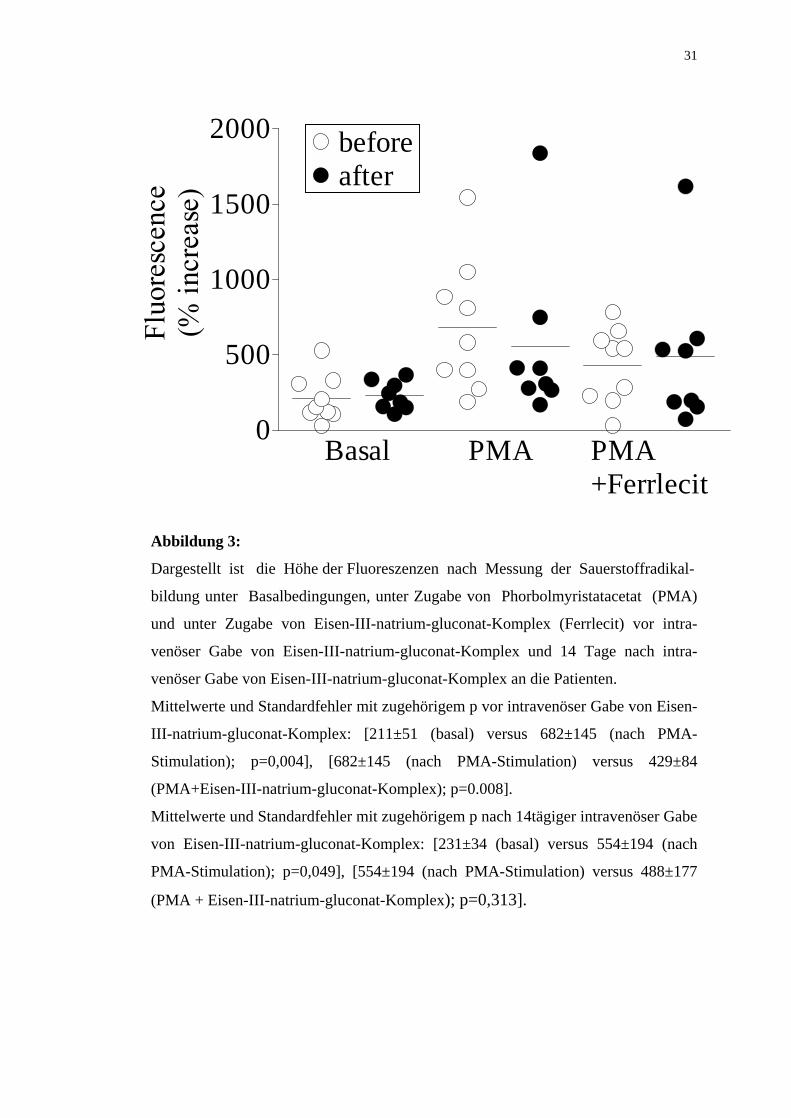
Abbildung 3:
Dargestellt ist die Höhe der Fluoreszenzen nach Messung der Sauerstoffradikal-
bildung unter Basalbedingungen, unter Zugabe von Phorbolmyristatacetat (PMA)
und unter Zugabe von Eisen-III-natrium-gluconat-Komplex (Ferrlecit) vor intra-
venöser Gabe von Eisen-III-natrium-gluconat-Komplex und 14 Tage nach intra-
venöser Gabe von Eisen-III-natrium-gluconat-Komplex an die Patienten.
Mittelwerte und Standardfehler mit zugehörigem p vor intravenöser Gabe von Eisen-
III-natrium-gluconat-Komplex: [211±51 (basal) versus 682±145 (nach PMA-
Stimulation); p=0,004], [682±145 (nach PMA-Stimulation) versus 429±84
(PMA+Eisen-III-natrium-gluconat-Komplex); p=0.008].
Mittelwerte und Standardfehler mit zugehörigem p nach 14tägiger intravenöser Gabe
von Eisen-III-natrium-gluconat-Komplex: [231±34 (basal) versus 554±194 (nach
PMA-Stimulation); p=0,049], [554±194 (nach PMA-Stimulation) versus 488±177
(PMA + Eisen-III-natrium-gluconat-Komplex); p=0,313].
0
500
1000
1500
2000
Basal PMA PMA+Ferrlecit
beforeafter
Page 32
32
Eisen-III-natrium-gluconat-Komplex konnte die PMA-induzierte Sauerstoff-
radikalbildung zwei Wochen nach intravenöser Gabe von Eisen-III-natrium-
gluconat-Komplex im Vergleich zur Verminderung der Produktion von Sauer-
stoffradikalen zwei Wochen vor intravenöser Gabe von Eisen-III-natrium-gluconat-
Komplex an die Patienten nicht zusätzlich senken (siehe hierzu Abbildung 3).
Jetzt wurden die Effekte von Eisen-III-natrium-gluconat-Komplex (Ferrlecit),
auf die Sauerstoffradikalproduktion von mononukleären Leukozyten in vitro unter-
sucht. Dazu wurden die reaktiven Sauerstoffradikale mit dem Fluoreszenz-Farbstoff
Dichlorofluoreszindiacetat, DCFDA, unter Basalbedingungen, 30 Minuten und 60
Minuten nach Vorgabe des Stimulators der Protein-Kinase-C, Phorbolmyristatacetat,
gemessen.
Zur Erstellung einer Dosiswirkungskurve für Eisen-III-natrium-gluconat-Komplex
wurde die Sauerstoffradikalbildung in mononukleären Leukozyten von Patienten mit
terminaler Niereninsuffizienz nach Stimulation mit PMA und nach Zugabe von
jeweils 1µl, 5µl, 10µl, 20µl, 50µl und 100µl Eisen-III-natrium-gluconat-Komplex
nach 0, 30 und 60 Minuten gemessen. Die Dosiswirkungskurve ist in der Abbildung
4 dargestellt. Mit zunehmender Konzentration an Eisen-III-natrium-gluconat-
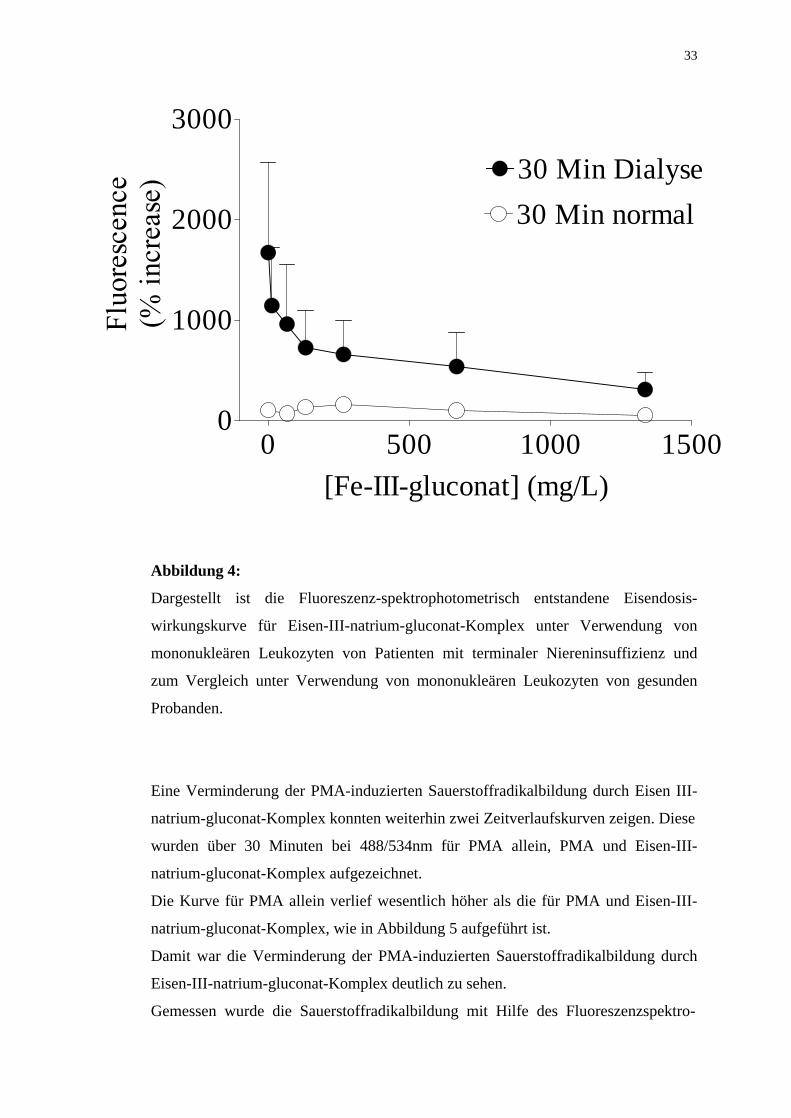
Komplex nahm die PMA-induzierte Sauerstoffradikalbildung in mononukleären
Zellen von Dialysepatienten von 1673±892 % auf 310±174 % (n=3; p=0,199)
signifikant ab. Dieser Effekt war insbesondere nach 30 Minuten ausgeprägt
Damit zeigte sich die Wirkung von Eisen-III-natrium-gluconat-Komplex sowohl
dosis- als auch zeitabhängig.
Zum Vergleich wurde der Versuch der Erstellung einer Dosiswirkungskurve für
Eisen-III-natrium-gluconat-Komplex unter Zuhilfenahme von mononukleären
Leukozyten von drei gesunden Probanden unternommen. In diesem Fall konnte
Eisen-III-natrium-gluconat-Komplex jedoch die Produktion von reaktiven Sauer-
stoffradikalen nicht signifikant verändern.
Die Abbildung 4 zeigt eine signifikante Verminderung der PMA-induzierten
Sauerstoffradikalbildung mit zunehmender Konzentration an Eisen-III-natrium-
gluconat-Komplex (1µl, 5µl, 10µl, 20µl, 50µl und 100µl). Dieser Effekt zeigte
sich nur in mononukleären Leukozyten von Patienten mit terminaler
Niereninsuffizienz, nicht jedoch in Leukozyten von gesunden Probanden.
Page 33
33
Abbildung 4:
Dargestellt ist die Fluoreszenz-spektrophotometrisch entstandene Eisendosis-
wirkungskurve für Eisen-III-natrium-gluconat-Komplex unter Verwendung von
mononukleären Leukozyten von Patienten mit terminaler Niereninsuffizienz und
zum Vergleich unter Verwendung von mononukleären Leukozyten von gesunden
Probanden.
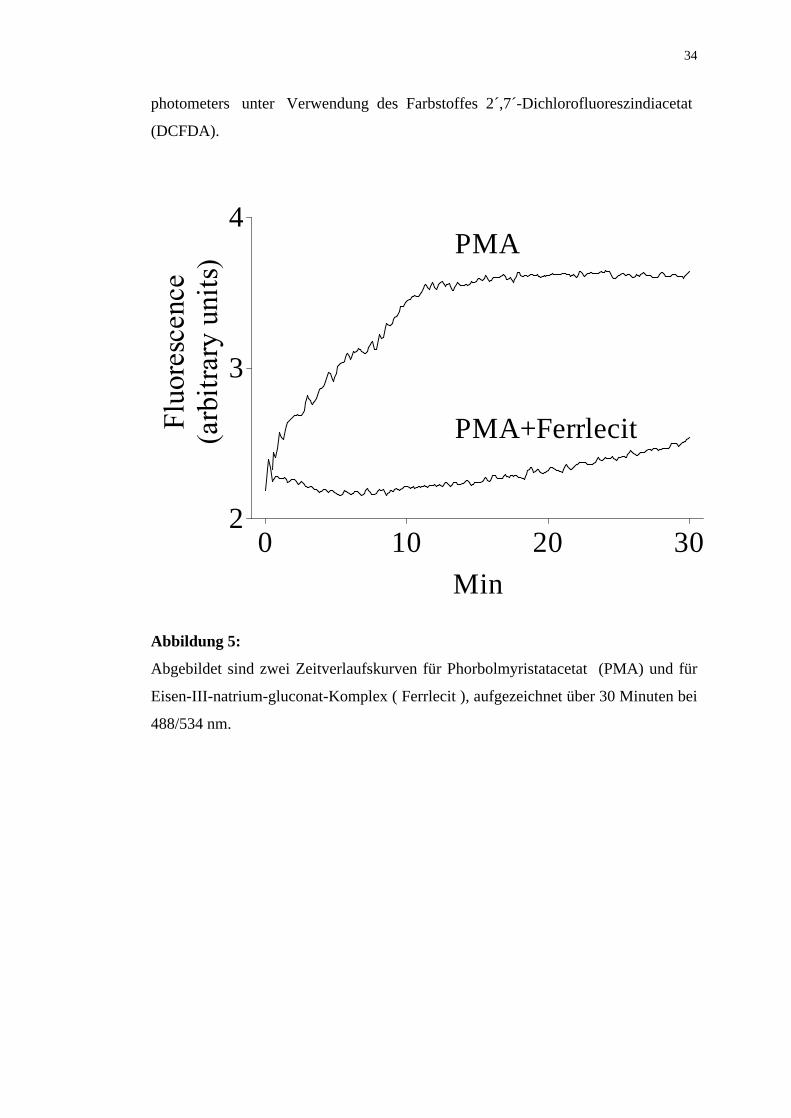
Eine Verminderung der PMA-induzierten Sauerstoffradikalbildung durch Eisen III-
natrium-gluconat-Komplex konnten weiterhin zwei Zeitverlaufskurven zeigen. Diese
wurden über 30 Minuten bei 488/534nm für PMA allein, PMA und Eisen-III-
natrium-gluconat-Komplex aufgezeichnet.
Die Kurve für PMA allein verlief wesentlich höher als die für PMA und Eisen-III-
natrium-gluconat-Komplex, wie in Abbildung 5 aufgeführt ist.
Damit war die Verminderung der PMA-induzierten Sauerstoffradikalbildung durch
Eisen-III-natrium-gluconat-Komplex deutlich zu sehen.
Gemessen wurde die Sauerstoffradikalbildung mit Hilfe des Fluoreszenzspektro-
0 500 1000 15000
1000
2000
3000
30 Min normal
30 Min Dialyse
[Fe-III-gluconat] (mg/L)
Page 34
34
photometers unter Verwendung des Farbstoffes 2´,7´-Dichlorofluoreszindiacetat
(DCFDA).
Abbildung 5:
Abgebildet sind zwei Zeitverlaufskurven für Phorbolmyristatacetat (PMA) und für
Eisen-III-natrium-gluconat-Komplex ( Ferrlecit ), aufgezeichnet über 30 Minuten bei
488/534 nm.
0 10 20 302
3
4PMA
PMA+Ferrlecit
Min
Page 35
35
5.2 WIRKUNG VON VANADATE, DISULFIRAM, DOG UND
EISEN-III-GLUCONAT
Vanadate, ein Hemmer von ATPasen, Phosphotransferasen-und kinasen, stimuliert
die NADP-Oxidase und begünstigt die Sauerstoffradikalbildung (Kalyani et al.,
1992; Shiel et al., 1992; Vijaya et al., 1984).
In Kombination mit PMA konnte die Fluoreszenz geringfügig gesteigert werden.
Die Vorgabe von Vanadate zu PMA führte zu einer Veränderung der PMA-
induzierten Sauerstoffradikalbildung von 909±161 % (n=26 bei Berücksichtigung
aller gemessenen Fluoreszenzwerte nach PMA-Stimulation) auf 1195±280 % (n 6;
p=0.218).
Nach Zugabe von 50µl Eisen-III-natrium-gluconat-Komplex zu PMA und zu PMA
zusammen mit Vanadate verminderte sich die Sauerstoffradikalbildung.
Die Vorgabe von Eisen-III-natrium-gluconat-Komplex führte gegenüber der PMA-
stimulierten Bildung von Sauerstoffradikalen zu einer Verminderung der Produktion
von Sauerstoffradikalen von 909±161 % auf 482±87 % (n 17; p=0.050).
Die Kombination PMA, 10µl Vanadate und 50µl Eisen-III-gluconat ergab gegenüber
der alleinigen Gabe von PMA ebenfalls eine Senkung der Sauerstoffradikalbildung
von 909±161 % auf 738±184 % (n 7; p=0.605).
Disulfiram, ein Inhibitor der Alkohol-und Aldehyddehydrogenase I+II und der
Caspase-1 und Caspase-3, gilt als Sauerstoffradikalfänger (Zhao et al., 2000).
Die Vergabe von Disulfiram nach PMA-induzierter Sauerstoffradikalbildung führte
entsprechend zu einer Senkung der Bildung von Sauerstoffradikalen von 909±161 %
auf 323±113 % (n=8; p=0.002).
Die Zugabe von 50µl Eisen-III-natrium-gluconat-Komplex zu PMA und Disulfiram
konnte die Sauerstoffradikalbildung gegenüber der alleinigen Gabe von PMA von
909±161 % auf 207±55 % senken (n 7; p=0.008).
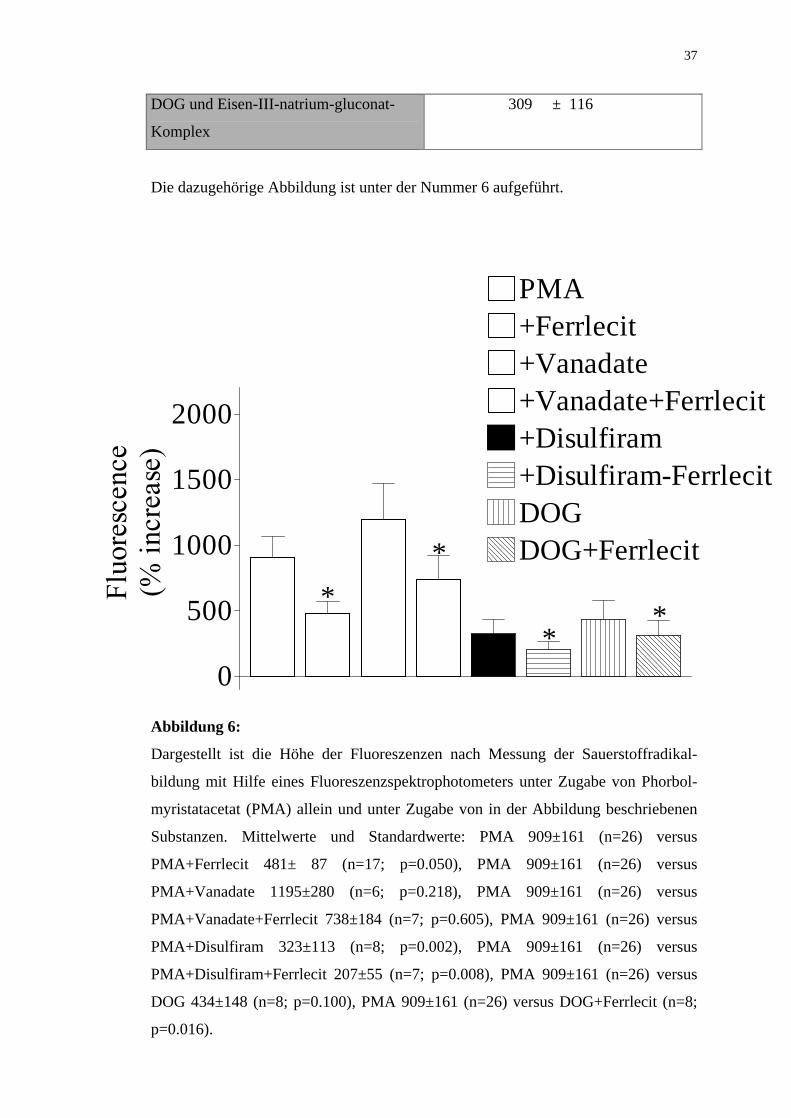
Der Fluoreszenz-mindernde Effekt von Eisen-III-natrium-gluconat-Komplex ist
somit durch Vanadate nicht aufhebbar.
Zur Prüfung, ob neben PMA auch weitere Stimulatoren der Proteinkinase C eine
Steigerung der Bildung von reaktiven Sauerstoffradikalen zeigen, wurde DOG, 1,2
Dioctanoyl-sn-glycerol, eingesetzt. Die Zugabe von DOG führte zu einer Bildung
von reaktiven Sauerstoffradikalen wie in Studien zuvor beschrieben (Follin et al.,
Page 36
36
1991), allerdings in geringerem Maße als bei Zugabe von PMA. Die Fluoreszenz
betrug 434±148 % (n=8; p=0,100) versus 909±161 % nach alleiniger Gabe von PMA
(n 26).
Nach Vergabe von Eisen-III-natrium-gluconat-Komplex zu DOG betrug die Bildung
von reaktiven Sauerstoffradikalen 309±116 % (n 8) versus 909±161 % (n=26) nach
alleiniger Gabe von PMA (p= 0.016).
Die Mittelwerte der gemessenen Fluoreszenzen nach Zugabe von PMA, Eisen-III-
natrium-gluconat-Komplex, PMA und Vanadate, PMA, Vanadate und Eisen-III-
natrium-gluconat-Komplex, PMA und Disulfiram, PMA, Disulfiram und Eisen-III-
natrium-gluconat-Komplex, DOG allein, DOG und Eisen-III-natrium-gluconat-
Komplex sind in der Tabelle 3 aufgeführt.
Auffällig ist, daß die Wirkung von Eisen-III-natrium-gluconat-Komplex (Ferrlecit)
weder durch Vanadate noch durch Disulfiram aufhebbar war.
Tabelle 3:
Angegeben sind die prozentualen Veränderungen der gemessenen Fluoreszenzwerte
nach Stimulation mit Phorbolmyristatacetat (PMA) und nach Zugabe der in der
Tabelle aufgeführten Substanzen als Mittelwerte Standardfehler:
PMA 909 ± 161
PMA und Vanadate 1195 ± 280
PMA und Eisen-III-natrium-gluconat-
Komplex
482 ± 87
PMA, Vanadate und Eisen-III-natrium-
gluconat-Komplex
738 ± 184
PMA und Disulfiram 323 ± 113
PMA, Disulfiram und Eisen-III-natrium
gluconat-Komplex
207 ± 55
DOG 434 ± 148
Page 37
37
DOG und Eisen-III-natrium-gluconat-
Komplex
309 ± 116
Die dazugehörige Abbildung ist unter der Nummer 6 aufgeführt.
Abbildung 6:
Dargestellt ist die Höhe der Fluoreszenzen nach Messung der Sauerstoffradikal-
bildung mit Hilfe eines Fluoreszenzspektrophotometers unter Zugabe von Phorbol-
myristatacetat (PMA) allein und unter Zugabe von in der Abbildung beschriebenen
Substanzen. Mittelwerte und Standardwerte: PMA 909±161 (n=26) versus
PMA+Ferrlecit 481± 87 (n=17; p=0.050), PMA 909±161 (n=26) versus
PMA+Vanadate 1195±280 (n=6; p=0.218), PMA 909±161 (n=26) versus
PMA+Vanadate+Ferrlecit 738±184 (n=7; p=0.605), PMA 909±161 (n=26) versus
PMA+Disulfiram 323±113 (n=8; p=0.002), PMA 909±161 (n=26) versus
PMA+Disulfiram+Ferrlecit 207±55 (n=7; p=0.008), PMA 909±161 (n=26) versus
DOG 434±148 (n=8; p=0.100), PMA 909±161 (n=26) versus DOG+Ferrlecit (n=8;
p=0.016).
0
500
1000
1500
2000
PMA+Ferrlecit+Vanadate+Vanadate+Ferrlecit+Disulfiram+Disulfiram-FerrlecitDOGDOG+Ferrlecit
**
*
*
Page 38
38
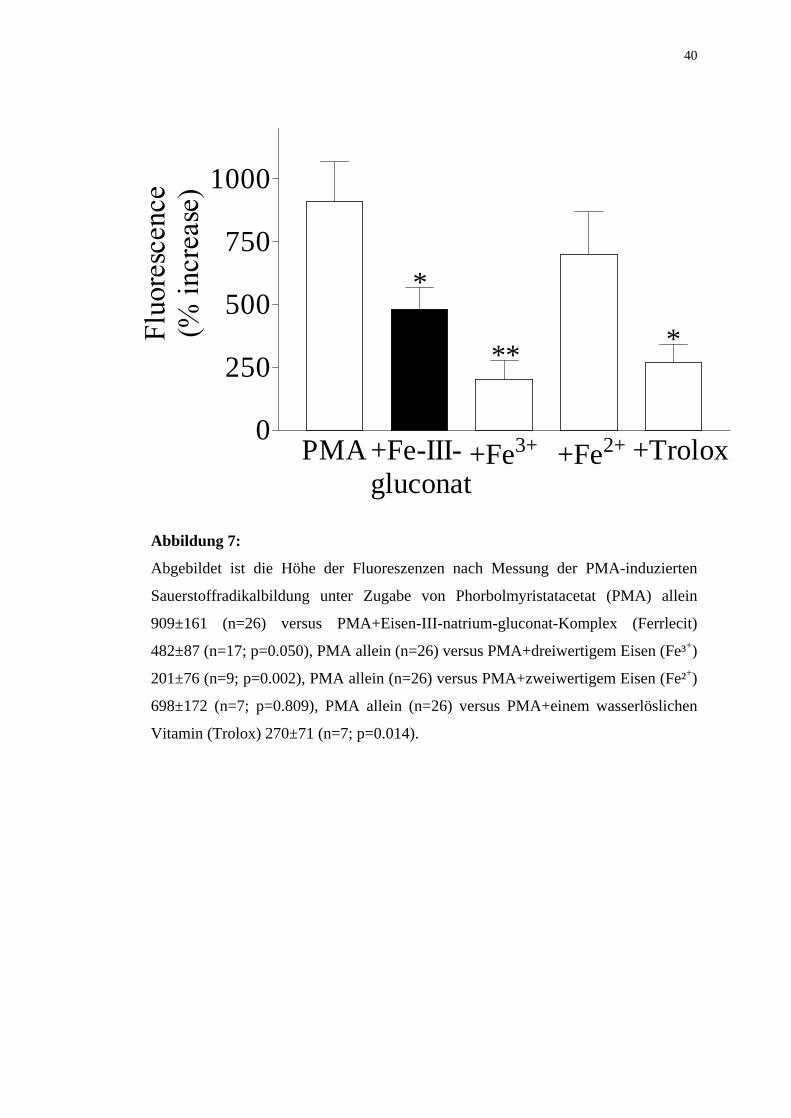
5.3 WIRKUNG VON FeCl3, FeCl2 UND TROLOX
Ziel dieses Versuchs war die Mechanismusaufklärung bezüglich der Wirkung von
Eisen-III-natrium-gluconat-Komplex.
Der Eisen-III-natrium-gluconat-Komplex vermochte in den genannten Versuchen die
PMA-induzierte Sauerstoffradikalbildung zu hemmen.
Nach Zugabe von 50µl dreiwertigem Eisen, FeCl3, zu 10µl PMA sank die PMA-
induzierte Sauerstoffradikalbildung von 909±161 % (n=26) auf 201±76 % (n 9;
p=0.002).
Die Zugabe von 50µl zweiwertigem Eisen, FeCl2, vermochte die PMA-induzierte
Sauerstoffradikalbildung nur geringfügig zu senken.
Die Zugabe von 50µl FeCl2 führte zu einer Verminderung der PMA-induzierten
Bildung von reaktiven Sauerstoffradikalen von 909±161 % (n=26) auf 698±172 %
(n 7; p=0.807).
Zur Kontrolle wurde PMA, Trolox, 6-Hydroxyy-2,5,7,8-tetramethylchroman-2-
carboxylsäure, zugesetzt. Das wasserlösliche Vitamin E-Derivat gilt als antioxidativ
(Roob et al., 2000; Uysal et al., 1998). Die Sauerstoffradikalbildung sank nach
Zugabe von Trolox signifikant von 909±161 % (n=26) auf 270±71 % (n 7;p=0.014).
Die Mittelwerte der Fluoreszenzen und deren Prozentzahlen nach Zugabe von
Trolox, FeCl2 und FeCl3 sind in der Tabelle 4 zu finden.
Die Abbildung 7 veranschaulicht die Verminderung bzw. die Steigerung der
Sauerstoffradikalbildung nach Zugabe von Trolox, FeCl2 und FeCl3.
Die Sauerstoffradikalbildung wurde Fluoreszenz-spektrophotometrisch mit dem
Farbstoff 2´,7´-Dichlorofluoreszindiacetat (DCFDA) gemessen.
Auffällig war die signifikante Verminderung der PMA-induzierten Sauerstoff-
radikalbildung durch Eisen-III-natrium-gluconat-Komplex, noch signifikanter durch
Fe³+ und durch Trolox, nicht jedoch durch Fe²+.
Page 39
39
Tabelle 4:
Angegeben sind die prozentualen Veränderungen der gemessenen Fluoreszenzwerte
nach Stimulation mit Phorbolmyristatacetat (PMA) und unter Zugabe von drei-
wertigem Eisenchlorid (FeCl3) und zweiwertigem Eisenchlorid (FeCl2) als
Mittelwerte Standardfehler:
PMA 909 ± 161
PMA und FeCl3
201 ± 76
PMA und FeCl2
698 ± 172
PMA und Trolox
270 ± 71
Page 40
40
Abbildung 7:
Abgebildet ist die Höhe der Fluoreszenzen nach Messung der PMA-induzierten
Sauerstoffradikalbildung unter Zugabe von Phorbolmyristatacetat (PMA) allein
909±161 (n=26) versus PMA+Eisen-III-natrium-gluconat-Komplex (Ferrlecit)
482±87 (n=17; p=0.050), PMA allein (n=26) versus PMA+dreiwertigem Eisen (Fe³+)
201±76 (n=9; p=0.002), PMA allein (n=26) versus PMA+zweiwertigem Eisen (Fe²+)
698±172 (n=7; p=0.809), PMA allein (n=26) versus PMA+einem wasserlöslichen
Vitamin (Trolox) 270±71 (n=7; p=0.014).
0
250
500
750
1000
PMA +Fe-III-gluconat
+Fe3+ +Fe2+
*
**
+Trolox
*
Page 41
41
5.4 VERSUCHE MIT DEM EISENINDIKATOR PHEN GREEN
Der Farbstoff phen green ist in der Lage zweiwertiges Eisen, FeCl2, zu binden und
damit die Anwesenheit von zweiwertigem Eisen zu messen (Petrat et al., 2000).
Nach Inkubation mit 10µmol/L phen green, wurden den sich in den Küvetten
befindlichen mononukleären Leukozyten zur Eichung Digitonin und FeCl2 zugesetzt.
Der Anstieg von zweiwertigem Eisen wurde unter Basalbedingungen und nach einer
Inkubation von Eisen-III-natrium-gluconat-Komplex (Ferrlecit) nach 30 und 60
Minuten ermittelt.
Nach einer Inkubation mit Eisen-III-natrium-gluconat-Komplex von 30 Minuten
stieg zweiwertiges Eisen von 77 5 % auf 85 6 % an (n 7; p=0.016). Die Inkubation
mit Eisen-III-natrium-gluconat-Komplex von 60 Minuten führte zu einem Anstieg an
zweiwertigem Eisen von 77 5 % auf 91 6 % ( n 7; p=0.031).
Die ermittelten Prozentzahlen sind in der Tabelle 5 zu finden.
Tabelle 5:
Angegeben sind die prozentualen Veränderungen der gemessenen Fluoreszenzwerte
als Mittelwerte
Standardfehler nach Messung mit dem Farbstoff phen green unter
Kontrollbedingungen und nach einer Inkubation mit Eisen-III-natrium-gluconat-
Komplex (Ferrlecit) nach 30 und 60 Minuten.
Kontrolle Inkubation mit Eisen-III-
gluconat nach 30
Minuten
Inkubation mit Eisen-III-
gluconat nach 60
Minuten
77 ± 5
85 ± 6
91 ± 6
Die Abbildungen 8, 9 und 10 zeigen die Höhe der Fluoreszenzen unter Verwendung
des Farbstoffes phen green, von Eisen-III-natrium-gluconat-Komplex, von
Eisenchlorid und unter Verwendung von Nigericin und Digitonin.
Page 42
42
Die Fluoreszenzen wurden Fluoreszenz-spektrophotometrisch bestimmt. Der Farb-
stoff phen green bindet zweiwertiges Eisen und damit kommt es zu einer Abnahme
der Fluoreszenz.
Die Abbildungen 8 und 10 zeigen, daß die Inkubation mit Eisen-III-natrium-
gluconat-Komplex sowohl nach 30 als auch nach 60 Minuten zweiwertiges Eisen
erhöhte.
In der Abbildung 9 wird verdeutlicht, daß die Zugabe von Nigericin und
zweiwertigem Eisenchlorid (FeCl2) zu einer Erhöhung an zweiwertigem Eisen
führte, während die Zugabe von Digitonin zweiwertiges Eisen verminderte.
Abbildung 8:
Dargestellt ist die Höhe der Fluoreszenzen nach Messung mit einem
Fluoreszenzspektrophotometer unter Verwendung des Farbstoffes phen green. Die
Fluoreszenzen wurden unter Basalbedingungen und unter Inkubation mit Eisen-III-
natrium-gluconat-Komplex nach 30 und 60 Minuten gemessen.
60
80
100
basalFe-III-gluconat
*
*
30 Min 60 Min FeCl2
Page 43
43
Abbildung 9:
Dargestellt ist eine Kurve, die unter Zugabe von Nigericin, einem Antibiotikum,
Digitonin und unter Zugabe von zweiwertigem Eisenchlorid (FeCl2) über 400
Sekunden aufgezeichnet wurde.
0
1000
2000Nigericin
Digitonin
FeCl2
Control
100s
Page 44
44
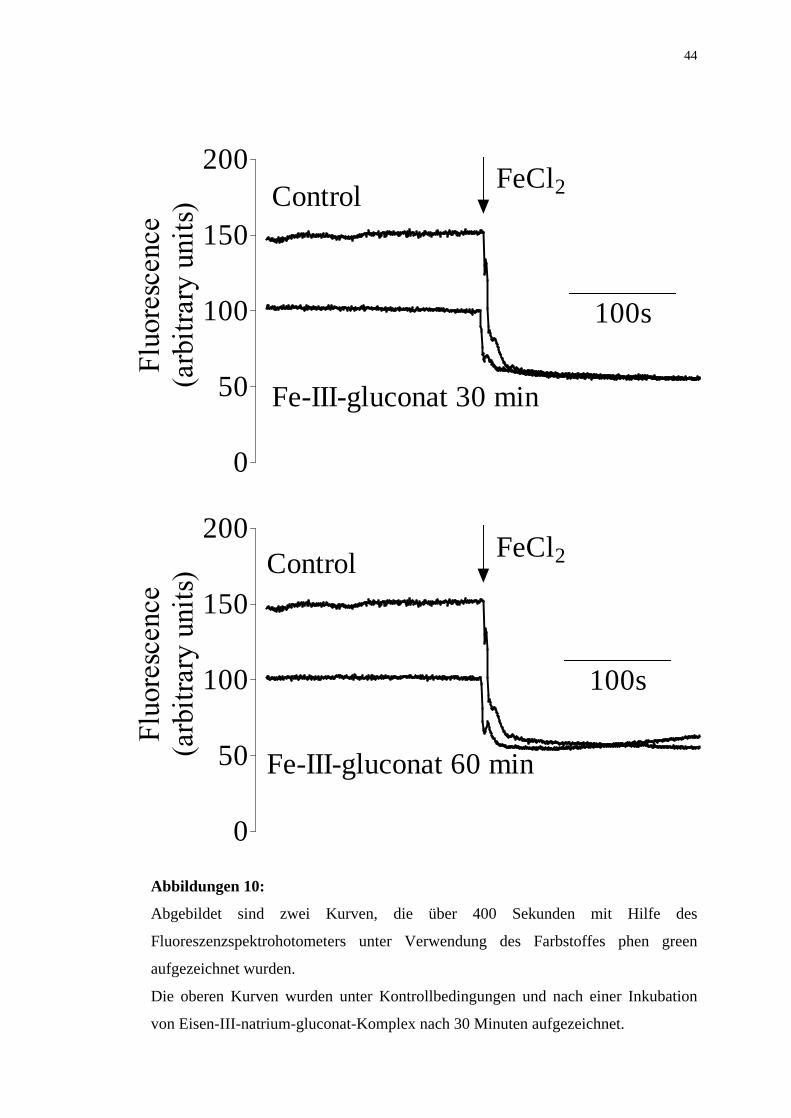
Abbildungen 10:
Abgebildet sind zwei Kurven, die über 400 Sekunden mit Hilfe des
Fluoreszenzspektrohotometers unter Verwendung des Farbstoffes phen green
aufgezeichnet wurden.
Die oberen Kurven wurden unter Kontrollbedingungen und nach einer Inkubation
von Eisen-III-natrium-gluconat-Komplex nach 30 Minuten aufgezeichnet.
0
50
100
150
200FeCl2
100s
Control
Fe-III-gluconat 30 min
0
50
100
150
200FeCl2
100s
Control
Fe-III-gluconat 60 min
Page 45
45
Die unteren Kurven wurden ebenfalls unter Kontrollbedingungen und nach einer
Inkubation von Eisen-III-natrium-gluconat-Komplex nach 60 Minuten erfasst.
Dieser Versuch mit dem Fluoreszenz-Marker für intrazelluläres Eisen, phen green,
zeigt, daß die Inkubation der mononukleären Leukozyten mit Eisen-III-natrium-
gluconat-Komplex das intrazelluläre Eisen-II erhöht.
Durch Eisen-III-natrium-gluconat-Komplex kommt es zu einer Steigerung von
intrazellulärem Eisen-II. Mittels der Fenton-Reaktion erfolgt eine verstärkte Bildung
von Eisen-III und Hydroxlyradikalen. Umgekehrt werden die durch den Fluoreszenz-
Farbstoff Dichlorofluoreszindiacet, DCFDA, detektierten Hydrogenperoxidradikale
(Bustamante et al., 1997) nach Eisen-III-gluconat intrazellulär vermindert.
5.5 KONTROLLVERSUCHE IN ABWESENHEIT VON
MONONUKLEÄREN LEUKOZYTEN
Eisen-III-natrium-gluconat-Komplex konnte in den zuvor beschriebenen Versuchen
in Anwesenheit von mononukleären Leukozyten die PMA-induzierte Sauerstoff-
radikalbildung hemmen, sofern die mononukleären Leukozyten von Dialysepatienten
stammten. Die Hemmung der Sauerstoffradikalbildung durch Eisen-III-natrium-
gluconat-Komplex resultiert dabei aus einer Wirkung auf die mononukleären
Leukozyten selbst. Um zu zeigen, daß die Verminderung der Fluoreszenz durch
Eisen-III-natrium-gluconat-Komplex nicht etwa auf einer Wirkung auf den Farbstoff
Dichlorofluoreszindiacetat, DCFDA, beruht, wurden Versuche in Abwesenheit von
mononukleären Leukozyten durchgeführt.
Es wurden 1000µl „Hank´s balanced salt solution“ ,HBSS, 10µl Eisen-III-natrium-
gluconat-Komplex und anschließend jeweils 1000µl HBSS oder 1000µl H2O2
zugesetzt. Durch Zugabe von HBSS oder H2O2 erfolgte jedoch keine Veränderung
der Fluoreszenz von Eisen-III-natrium-gluconat-Komplex.
Im weiteren Versuch wurden der pH-Wert und die Fluoreszenz von 1000µl HBSS
allein und anschließend unter Zugabe von zunächst 10µl DCFDA, dann 10µl Eisen-
Page 46
46
III-natrium-gluconat-Komplex, 40µl Eisen-III-natrium-gluconat-Komplex und unter
Zugabe von 50µl Eisen-III-natrium-gluconat-Komplex gemessen.
Die Fluoreszenz stieg unter Zugabe von DCFDA, blieb aber unter weiterer Zugabe
von Eisen-III-natrium-gluconat-Komplex nahezu unverändert. Der pH-Wert sank
von 7,32 auf 7,12 nach Zugabe des Farbstoffes DCFDA, blieb aber unter weiterer
Zugabe von Eisen-III-natrium-gluconat-Komplex konstant bei 7,14.
Damit konnte gezeigt werden, daß Eisen-III-natrium-gluconat-Komplex keinen
Einfluß auf den Farbstoff DCFDA und ebenfalls keinen Einfluß auf den pH-Wert
hatte.
Page 47
47
5. DISKUSSION
Die Fragestellung dieser Arbeit ist die Wirkung von Eisen-III-natrium-gluconat-
Komplex (Ferrlecit) auf die Sauerstoffradikalbildung bei Patienten mit terminaler
Niereninsuffizienz, die eine Dialysetherapie erhalten.
Eisen-III-natrium-gluconat-Komplex erhalten Patienten mit chronischer Nieren-
insuffizienz nach der Dialyse neben humanem, rekombinantem Erythropoetin.
Patienten mit terminaler Niereninsuffizienz leiden unter einer normozytären Anämie,
überwiegend als Folge eines Erythropoetinmangels. Erythropoetin ist ein 34-39 kD
großes Glykoprotein und wird in peritubulären Zellen in der Niere sowie in der
fetalen Leber gebildet. Erythropoetin stimuliert Wachstum und Differenzierung von
frühen erythropoetischen Vorläuferzellen. Außerdem induziert es zusammen mit
anderen Faktoren die Proliferation von Megakaryozyten-Vorläuferzellen. Die
chronische Niereninsuffizienz führt in der Regel zu einer verminderten Erythropoese,
die sich ab einem Serum-Kreatininwert von 3,5mg/dl als renale Anämie zeigt.
Hauptursache der renalen Anämie ist die Unfähigkeit der erkrankten Nieren, den
Hämoglobinspiegel durch eine adäquate Erythropoetinproduktion im Normbereich
zu halten. Die Serum-Erythropoetinspiegel liegen zwar bei Dialysepatienten meist im
Normbereich, sind aber inadäquat im Vergleich zu nierengesunden, gleichermaßen
anämischen Patienten, deren Werte um das 10-100fache höher liegen. Zusätzlich zur
Erythrozytenbildungsstörung besteht bei chronischer Niereninsuffizienz eine
negative Erythrozytenbilanz durch Blutverlust während der Dialyse und durch
gehäufte gastrointestinale Blutungen infolge urämischer Thrombozytopathie sowie
durch eine mäßige Verkürzung der Erythrozytenüberlebenszeit.
Seit es 1985 gelungen ist, das humane Gen, das Erythropoetin kodiert, zu
identifizieren, lassen sich größere Mengen des Hormons gentechnisch gewinnen
(rekombinantes, humanes Erythropoetin). Damit ist es möglich geworden, die renale
Anämie kausal und effektiv durch die Gabe von Erythropoetin zu korrigieren. Auf
die früher notwendigen problematischen Bluttransfusionen kann deshalb heute in
vielen Fällen verzichtet werden (Classen, 1994).
Eine weitere Ursache der renalen Anämie bei Patienten mit chronischer
Niereninsuffizienz stellt der Eisenmangel dar (Silverberg et al., 1999).
Page 48
48
Eisen, das in Hämoglobin enthalten ist, ist am Sauerstofftransport und als Kofaktor
in vielen enzymatischen Systemen beteiligt (Granick, 1958).
Eisen ist im retikuloendothelialen System der Leber, der Milz und des
Knochenmarks gespeichert. Ferner ist es an Ferritin und Hämosiderin gebunden.
Diese Eisenspeicher machen dreiviertel von 3 bis 5 g des totalen Eisenbestandes des
Körpers aus. Das restliche Eisen ist im erythropoetischem Gewebe oder in roten
Blutkörperchen enthalten. Das zirkulierende an Transferrin gebundene Eisen macht
nur 3 bis 4 mg des geamten Eisenbestandes aus. Die Eisenzufuhr kann bis zu 200 mg
bei Patienten mit terminaler Niereninsuffizienz betragen.
Allerdings, werden nur 1% bis 2% dieses Eisens resorbiert (Kooistra et al., 1995).
Der Eisenaustausch zwischen dem roten Knochenmark, den zirkulierenden roten
Blutkörperchen und zwischen dem retikuloendothelialen System beträgt 20 mg oder
mehr täglich.
Studien, die Knochenmark bei anämischen Patienten mit chronischer Nierenin-
suffizienz untersuchten, weisen auf verminderte Eisenspeicher trotz normalen
Serumeisens bei diesen Patienten hin (Goltloib et al., 1983).
Zusätzlich zu einem erhöhten Bedarf an Eisen bei gleichzeitiger Therapie mit
Erythropoetin, werden drei Mechanismen, die einen Eisenmangel bei Dialyse-
patienten bedingen, vorgeschlagen. Dazu gehören eine verminderte Eisenresorption,
externer Blutverlust und ein funktioneller Eisenmangel (Fishbane et al., 1997).
Eisen wird vor allem im Duodenum und im proximalen Jejunum resorbiert. Die
Resorption ist abhängig von der alimentären Eisenzufuhr, intraluminalen Faktoren,
der Aktivität der Erythropoese, der funktionellen Kapazität von intestinalen
Mukosazellen und von der Größe des Eisenspeichers im Gewebe (Charlton et al.,
1983). Aufgenommenes Eisen wird zu der dreiwertigen Form reduziert und an
hochmolekulare Komplexe gebunden.
Dialysepatienten erhalten üblicherweise eine eiweißarme Diät, die die Reduktion des
tierischen Eiweißverlustes vorsieht. Die Reduktion von tierischem Eisen in der
Nahrung kann nicht nur zu einem prozentualen Abfall der Eisenzufuhr, sondern auch
zu einem prozentualen Abfall in der Eisenresorption führen. Es wird mehr Eisen, das
in tierischem Eiweiß enthalten ist, aufgenommen als das, das vom pflanzlichen
Eiweiß stammt (Silverberg et al., 1999). Zwar ist die Eisenresorption bei Individuen
mit normaler renaler Funktion mit 1 mg Eisen pro Tag ebenfalls niedrig, allerdings
Page 49
49
steigt die Eisenresorption mit erhöhter Erythropoese und einem Abfall an
intrazellulärem Eisenspeicher an.
Ein weiterer Grund für die verminderte Eisenresorption sind die Veränderungen der
intestinalen Mukosa als Folge einer Urämie. Eine Urämie führt zu histologischen
Veränderungen der intestinalen Mukosa, wie z.B zu einem vermehrten Untergang
von Krypten, einer Ansammlung von Entzündungszellen, einer Abnahme der
Aktivität von Dipeptidasen und zu einer Zunahme der Aktivität von Disaccharidasen.
Weiterhin können Calcium - und Aluminiumphosphat-Binder sowie H2-Rezeptor-
Antagonisten, die häufiger bei Patienten mit terminaler Niereninsuffizienz eingesetzt
werden, zu einer verminderten Eisenresorption führen.
Viele Faktoren, wie retiniertes Blut im Dialysator, häufige Blutentnahmen, okkulte
gastrointestinale Blutungen und Blutverluste nach Anlage und Wechsel von
Dialyseshunts führen zu verstärkten Blutverlusten bei Dialysepatienten.
Die häufigsten Läsionen, die bei Dialysepatienten zu einer gastrointestinalen Blutung
führen, sind erosive Gastritis oder duodenale Ulzera (Silverberg et al., 1999).
Die „Anemia Work Group of the Dialysis Outcomes Quality Initiative“ weist daruf
hin, daß 25-100 mg Eisen wöchentlich nötig wären, um die beschriebenen
Blutverluste auszugleichen (NKF-DOQI., 1997).
Der funktionelle Eisenmangel entsteht, wenn die üblichen Marker für den
Eisenbestand des Körpers keinen absoluten Eisenmangel angeben (Ferritin mehr als
100 ng/ml; Sättigung des Transferrins mit Eisen mehr als 20 %).
Allerdings reagieren Dialysepatienten auf eine Eisensubstitution mit einer Erhöhung
des Hämatokrits bei konstanter Erythropoetingabe.
Bei Patienten mit einem funktionellen Eisenmangel ist Eisen in nicht ausreichendem
Maße verfügbar, um dem vermehrten Bedarf an Eisen bei gleichzeitiger
Erythropoetintherapie gerecht zu werden (Cavill et al., 1992).
Bei einigen dieser Patienten ist die Unfähigkeit, Eisen schnell zu mobilisieren, auf
eine retikuloendotheliale Blockade (Means et al., 1992) zurückzuführen.
Dialysepatienten können unter okkulten Infektionen oder anderen Konditionen
leiden, die eine adäquate Antwort auf eine Eisentherapie und die Nutzung von
Eisenspeichern verhindern. Dieser Effekt könnte auf die Zunahme an zirkulierenden
Zytokinen hinweisen, die Makrophagen des retikuloendothelialen Systems anregen
mehr Eisen zu speichern und dieses nicht freizugeben.
Page 50
50
Der Effekt der Zytokine kann zu einem Abfall an endogener Erythropoetin-
produktion und einem verminderten Ansprechen auf endogenes oder exogenes
Erythropoetin führen (Johnson et al., 1990).
Diese Patienten haben typischerweise eine erhöhte Transferrinsättigung und ein
erhöhtes Serumferritin. Eine Eisentherapie führt bei diesen Patienten nicht zum
gewünschten Erfolg. Deshalb sollten sogenannte Responder oder Non-Responder auf
die Eisentherapie unter Kontrolle des Ferritins, der Transferrinsättigung und des
Hämatokrits erkannt werden.
Berücksichtigt werden sollte aber, daß Serumferritin ebenfalls ein Akut-Phase-
Protein darstellt und bei vielen chronischen Erkrankungen erhöht sein kann, obwohl
die Eisenspeicher niedrig sind.
Bei Patienten, die von einer Eisentherapie profitieren, hat sich Eisen-III-natrium-
gluconat-Komplex in Kombination mit Erythropoetin bei der Behandlung der renalen
Anämie als wirksam, sicher und nebenwirkungsärmer als mit alternierenden
Präparaten, wie Eisendextran beschrieben (Nissenson et al., 1999).
Als Nebenwirkungen von Eisen-III-natrium-gluconat-Komplex werden in seltenen
Fällen anaphylaktoide/anaphylaktische Reaktionen mit Ödemen an verschiedenen
Körperstellen, exanthematischen Hautveränderungen sowie Blutdruckabfälle
beschrieben. Im Vergleich zu Eisendextran treten die genannten Nebenwirkungen
bei Eisen-III-natrium-gluconat-Komplex viel seltener auf (Faich and Strobos, 1999).
Polysaccharide, wie Dextran, die in Eisen-III-natrium-gluconat-Komplex nicht
enthalten sind, gelten als die primären Verursacher von allergischen
Reaktionen des Typs I.
Während die orale Eisentherapie in vielen Fällen die renale Anämie nicht
entscheidend vermindern konnte, ist intravenöses Eisen und hier vor allem Eisen-III-
natrium-gluconat-Komplex in Kombination mit Erythropoetin in der Lage den
Hämatokrit zu erhöhen (Vogel S, 2000). Die Kombination von Erythropoetin und
Eisen-III-natrium-gluconat-Komplex hat einen additiven Effekt. Mit dem Einsatz
von Eisen-III-natrium-gluconat-Komplex kann die Dosis von Erythropoetin reduziert
und damit eine wichtige Nebenwirkung des Erythropoetins, wie die Erhöhung des
Blutdrucks minimiert werden.
Page 51
51
Sauerstoffradikale spielen für viele physiologische Prozesse, wie z.B für die
Infektabwehr eine wichtige Rolle. Eine von vielen physiologischen Antworten der
Leukozyten auf extrazelluläre Stimuli ist die Bildung dieser Sauerstoffradikale.
Allerdings ist ihre Überproduktion toxisch, übersteigt die Kapazität der zellulären
antioxidativen Abwehrmechanismen und wird für viele Erkrankungen, wie den
Diabetes mellitus, Atherosklerose, chronisch inflammatorische Erkrankungen, den
Alterungsprozess und nicht zuletzt für die Progression einer Niereninsuffizienz, unter
anderem bei Dialysepatienten, verantwortlich gemacht (Yoshioka et al., 1998).
Da freies Eisen als toxisch gilt (Baliga et al., 1997), die Haber Weiss Reaktion und
damit die Sauerstoffradikalbildung zu fördern vermag (Alfrey A, 1994), stellte sich
nun die Frage nach der Toxizität von Eisen-III-natrium-gluconat-Komplex, das die
renale Anämie bei Patienten mit terminaler Niereninsuffizienz in Kombination mit
Erythropoetin wirksam vermindern kann.
Die Messung der Sauerstoffradikalbildung nach Vergabe von Eisen-III-natrium-
gluconat-Komplex bei Patienten mit terminaler Niereninsuffizienz, die eine
Dialysetherapie erhalten, wurde jedoch bisher noch nicht durchgeführt und wurde
daher zum Ziel dieser Studie.
Die Messung der Sauerstoffradikalbildung mit Hilfe der Fluoreszenz-
spektrophotometrie zeigte, daß Eisen-III-natrium-gluconat-Komplex die PMA
induzierte Sauerstoffradikalbildung bei Patienten, die eine Dialysetherapie erhalten,
vermindert. Dieser Effekt zeigte sich nur bei Patienten, die eine terminale
Niereninsuffizienz bieten, nicht jedoch bei gesunden Probanden. Patienten mit
terminaler Niereninsuffizienz leiden generell unter einer erhöhten
Sauerstoffradikalproduktion (Canavese et al., 1987). Die Dialysetherapie vermag die
Produktion an Sauerstoffradikalen zusätzlich zu verstärken (Fiorillo et al., 1998).
Die Verminderung der PMA-induzierten Sauerstoffradikalbildung durch Eisen-III-
natrium-gluconat-Komplex zeigte sich nur in vitro. In vivo war dieser Effekt nicht
darstellbar.
In vitro führte die direkte Eisen-III-natrium-gluconat-Komplex-Administration zu
einer Hemmung der Sauerstoffradikabildung. Hingegen hatte die Gabe von Eisen-III-
natrium-gluconat-Komplex über 14 Tage bei den Patienten keinen senkenden Effekt
auf die Bildung von reaktiven Sauerstoffradikalen. Eine mögliche Erklärung für
Page 52
52
diesen Unterschied wäre z.B. die fortlaufende Neubildung der Blutzellen bei den
Patienten in vivo. Weiterhin ist die Konzentration an freiem Eisen in vivo aufgrund
der raschen Bindung an Transferrin deutlich geringer als bei den in vitro Versuchen.
Die Messung von Sauerstoffradikalen nach Gabe von Eisen-III-natrium-gluconat-
Komplex in Kombination mit Vanadate konnte die Verminderung der PMA-
induzierten Sauerstoffradikalbildung durch Eisen-III-natrium-gluconat-Komplex
nicht aufheben. Vanadate gilt als Stimulator der Sauerstoffradikalbildung (Kalyani et
al., 1992).
Versuche mit zwei-und dreiwertigem Eisen in Anwesenheit von mononukleären
Leukozyten haben gezeigt, daß nur dreiwertiges, jedoch nicht zweiwertiges Eisen die
PMA-induzierte Sauerstoffradikalbildung vermindert.
Dreiwertiges Eisen, das in Eisen-III-natrium-gluconat-Komplex enthalten ist, wird
reduziert und nimmt Elektronen auf:
Fe3+ + O2- —› Fe2+ + O2
Der Effekt von Eisen-III-natrium-gluconat-Komplex ist ferner zeit-und konzen-
trationsabhängig.
In vitro war der fluoreszenzmindernde Effekt von Eisen-III-natrium-gluconat-
Komplex vor allem nach 30 Minuten in einer Konzentration von 31,25µg/ml
besonders ausgeprägt.
Ob Heparin direkt die Bildung von Sauerstoffradikalen bei Patienten mit dialyse-
pflichtiger Niereninsuffizienz beeinflussen kann, ist nicht hinreichend bekannt.
Immerhin haben Dandona et al. beschireben, daß Heparin bei gesunden Probanden
die Bildung von reaktiven Sauerstoffradikalen vermindern kann. In der vorliegenden
Arbeit wurden die Messungen an Blutzellen durchgeführt, die aus heparinisiertem
Vollblut isoliert worden waren. Bei sonst gleichen Messbedingungen scheint die
Hemmung der Bildung von Sauerstoffradikalen durch Eisen-III-natrium-gluconat-
Komplex nicht durch den Heparin-Effekt bedingt zu sein.
Phorbolmyristatacetat, PMA, stimuliert die Proteinkinase C und in weiteren
Reaktionsschritten schließlich die NADPH-Oxidase, wodurch es zur Sauerstoff-
radikalbildung kommt. Dreiwertiges Eisen vermag Reaktionsschritte, die zur
Sauerstoffradikalbildung führen, zu verhindern bzw. zu mindern.
Page 53
53
Versuche in Abwesenheit von präparierten mononukleären Leukozyten haben
gezeigt, daß die fluoreszenzmindernde Wirkung von Eisen-III-natrium-gluconat-
Komplex auf einer Wirkung von Eisen-III-natrium-gluconat-Komplex auf die
mononukleären Leukozyten selbst beruht. In Abwesenheit von monunukleären
Leukozyten hatte Eisen-III-natrium-gluconat-Komplex keinen Einfluß auf die
Fluoreszenz und damit keinen Einfluß auf die Sauerstoffradikalbildung.
Da Eisen als potentiell schädlich wirkt und nach der Fenton-Reaktion die
Sauerstoffradikalbildung erhöht, stellte sich die Frage nach einer Aufklärung des
Mechanismus, der zu einer Verminderung der PMA-induzierten Sauerstoffradikal-
bildung durch Eisen-III-natrium-gluconat-Komplex führt. Zur Klärung dieser Frage
wurden Versuche mit dem Eisenindikator phen green durchgeführt. Phen green ist in
der Lage zweiwertiges Eisen zu binden und damit die Anwesenheit von FeCl2 zu
messen. Versuche mit dem Eisenindikator phen green haben gezeigt, daß eine
Inkubation mit Eisen-III-natrium-gluconat-Komplex zweiwertiges Eisen, FeCl2,
erhöht.
Unter Anwesenheit von Eisen-III-natrium-gluconat-Komplex kommt es zu einer
Erhöhung der Menge an zweiwertigem Eisen, FeCl2, und damit zu einer
Verschiebung des Gleichgewichtes in der Fenton-Reaktion auf die Seite Eisen-III +
OH*.
Die Fenton-Reaktion lautet: H2O2 + Fe II —> Fe III + OH*
Bei Stimulation mit PMA fällt immer die gleiche H2O2 - Menge an.
H2O2 wird von dem Farbstoff Dichlorofluoreszindiacetat, DCFDA, gemessen. So
steigt die DCF-Fluoreszenz linear mit der H2O2-Konzentration nach einer Inkubation
von 30 Minuten an (Bustamante., 1997).
DCFDA detektiert also vor allem H2O2, während OH*-Radikale von DCFDA
offenbar nicht gemessen werden.
Folglich führt die Anwesenheit von Eisen-III-natrium-gluconat-Komplex zu einer
Verminderung der PMA-induzierten Fluoreszenz und damit auch der PMA-
induzierten Sauerstoffradikalbildung.
Page 54
54
7. ZUSAMMENFASSUNG
Bekanntermaßen spielen reaktive Sauerstoffradikale eine wesentliche Rolle in der
Pathogenese unterschiedlicher Erkrankungen wie Atherosklerose, Niereninsuffizienz,
Tumoren oder Stoffwechselerkrankungen. In der vorliegenden Arbeit wurde der
Effekt von Eisen auf die Bildung von Sauerstoffradikalen in mononukleären
Leukozyten in vitro untersucht. Dazu wurden intrazelluläre reaktive
Sauerstoffradikale, insbesonders Hydrogenperoxid, mit Hilfe des Fluoreszenz-
Farbstoffes 2`,7`-Dichlorofluoreszindiacetat spektrophotometrisch bestimmt.
Die in-vitro Messungen wurden mit mononukleären Leukozyten von 16 Patienten
mit terminaler Niereninsuffizienz durchgeführt. Bei 9 Patienten mit terminaler
Niereninsuffizienz erfolgten die Messungen vor und 14 Tage nach Gabe von Eisen-
III-natrium-gluconat-Komplex.
Die basale Bildung von reaktiven Sauerstoffradikalen in mononukleären Leukozyten
war nicht signifikant unterschiedlich vor und 14 Tage nach Gabe von Eisen-III-
natrium-gluconat-komplex: 211±51 % versus 231±34 % nach 30 Minuten,
Mittelwerte ± Standardfehler des Mittelwertes, n=9.
In vitro konnte die Bildung von Sauerstoffradikalen durch den Stimulator der Protein
Kinase C, Phorbolmyristatacetat, durch Ebselen, Natriumazid, Captopril und Eisen-
III-natrium-gluconat-Komplex signifikant gehemmt werden. Die hemmende
Wirkung von Eisen-III-natrium-gluconat-Komplex auf die Bildung von reaktiven
Sauerstoffradikalen war zeit - und konzentrationsabhängig.
Vanadate ist als Stimulator der Sauerstoffradikalbildung bekannt. In der
Kombination mit Eisen-III-natrium-gluconat-Komplex konnte Vanadate die
Verminderung der Sauerstoffradikalbildung durch Eisen-III-natrium-gluconat-
Komplex nicht aufheben.
Versuche mit dem Fluoreszenz-Marker für intrazellulären Eisen, phen green, zeigten,
daß die Inkubation der mononukleären Leukozyten mit Eisen-III-natrium-gluconat-
Komplex das intrazelluläre Eisen-II erhöht.
Page 55
55
Die Untersuchung zeigt, daß es durch Eisen-III-natrium-gluconat-Komplex zu einer
Steigerung von intrazellulärem Eisen kommt. Mittels der Fenton-Reaktion erfolgt
eine verstärkte Bildung von Eisen-III und Hydroxylradikalen. Umgekehrt werden die
durch den Fluoreszenz-Farbstoff Dichlorofluoreszindiacetat detektierten Hydro-
genperoxidradikale durch Eisen-III-natrium-gluconat-komplex intrazellulär ver-
mindert.
Page 56
56
8. LITERATURVERZEICHNIS
1. Alfrey A
Role of iron and oxygen radicals in the progression of chronic
renal failure
Am J Kidney Dis 1994; 23: 183-187
2. Aruoma OI, Akanmu D, Cecchini R and Halliwell B
Evaluation of the ability of the angiotensin-converting enzyme inhibitor
Captpopril to scavenge reactive oxygen species
Chem Biol Inter 1991; 77: 303-314
3. Bailie G, Johnson C and Mason N
Parenteral iron use in the management of anemia in end-stage renal
disease patients
Am J. Kidney Dis 2000; 35: 5-11
4. Baliga R, Ueda N, Walker PD, Shah SV
Oxidant mechanism in toxic acute renal failure
Am J Kidney Dis 1997; 29: 465-477
5. Baum M and Hays SR
Phorbol myristate acetate and dioctanoylglycerol inhibit transport in
rabbit proximal convoluted tubule
Am Phys Soc 1998: 21: 120-124
6. Burns D and Pomposelli J
Toxicity of parenteral iron dextran therapy
Kidney Int 1999; 55: 119-124
7. Bustamante J, Tovar-B A, Montero G, Boveris A
Early redox changes during rat thymocyte apoptosis
Arch Biochem Biophys 1997; 337: 121-128
Page 57
57
8. Canavese C, Stratta P, Vercellone A
Oxygen free radicals in nephrology
Art Kidney and Dial 1987; 10: 379-389
9. Carmichael A
Reaction of vanadyl with hydrogen peroxide. An esr and spin
trapping study
Free Rad Res Comms 1995; 10: 37-45
10. Cavill I, Macdougall IC
Erythropoiesis and iron supply in patients treated with erythropoietin
Erythropoiesis 1992; 3: 50-55
11. Charlton RW, Bothwell TH
Iron absorption
Ann Rev Med 1983; 34: 55-68
12. Eschbach J and Adamson J
Iron overload in renal failure patients: Changes since the introduction of
erythropoietin therapy
Kidney Int 1999; 55: 35-43
13. Escobar JA, Rubio MA, and Lissi EA
SOD and catalase inactivation by singlet oxygen and peroxyl radicals
Free Rad Biol Med 1996; 20: 285-290
14. Evstigneva RP, Volkov IM and Chudinova VV
Vitamin E as a universal antioxidant and stabilizer of
biological membranes
Membr Cell Biol 1998; 12: 151-172
Page 58
58
15. Faich G and Strobos J
Sodium ferric gluconate complex in sucrose: safer intravenous iron
therapy than iron dextrans
Am J Kidney Dis 1999; 33: 464-70
16. Fiorillo C, Oliviero C, Rizzuti G, Nediani C, Pacini A, Nassi P.
Oxidative stress and antioxidant defenses in renal patients receiving regular
haemodialysis
Clin Chem Lab Med 1998; 36: 149-53
17. Fishbane S, Maesaka JK
Iron management in end-stage renal disease
Am J Kidney Dis 1997; 29: 319-333
18. Fishbane S, Mittal SK and Maesaka JK
Beneficial effects of iron therapy in renal failure patients
on hemodialysis
Kidney Int 1999; 69: 67-70
19. Follin P, Johansson A and Dahlgren C
Intracellular production of reactive oxygen species in human
neutrophils following activation by the soluble stimuli
FMLP, dioctanoylglycerol and ionomycin
Cell Biochem Funct 1991; 9: 29-37
20. Galli F, Canestrari F, Bellomo G.
Pathophysiology of the oxidative stress and ist implication in uremia and
dialysis
Contrib Nephrol 1999; 127: 1-31
21. Goltloib L, Ludicer H, Mines M, Ackermann J
Iron storage in chronic uremic undialyzed patients
Dial Transplant 1983; 12: 442-442
Page 59
59
22. Granick S
Iron metabolism
Bull NY Acad Med 1958; 30: 2-7
23. Gupta R, Gupta S, Joshi K and Ganguly NK
Role of iron and iron chelation therapy in oxygen free radical
mediated tissue injury in an ascending mouse model of chronic
pyelonehritis
Comp Immun Microbiol Infect Dis 1997; 20: 299-307
24. Halliwell B
Oxidants and human disease: some new concepts
FASEB J 1987; 1: 358-364
25. Hournant M
Le sndrome d´ischémie reperfusion rénale
Néphrologie 1999; 20: 371-375
26. Hultqvist M, Hebrant J, Nilsson-Thorell C, Lindholm T, Nilsson P,
Linden T and Hultqvist-Bengtsson U
Plasma concentration of vitamin C, vitamin E and/or malondialdehyde
as markers of oxygen free radical production during hemodialysis
Clin Nephrol 1997; 47: 37-46
27. Husain K, Morris C, Whitworth C, Trammell GL,
Rybak LP and Somani SM
Protection by ebselen against cisplatin-induced nephrotoxicity:
Antioxidant system
Depart Pharmacol Surg 1997: 21: 11-18
28. Johnson RA, Cook CA, Furmanski P
In vivo suppression of erythropoiesis by tumor necrosis factor-a
( TNF-alpha ): Reversal with exogenous erythropoietin
Exp Hematol 1990; 18: 109-115
Page 60
60
29. Jonas SK and Rileyn PA
Modification of the in vitro cytotoxicity of hydrogen peroxide by iron
complexes
Free Rad Res Comms 1995; 17: 407-419
30. Kaltwasser J and Gottschalk R
Erythropoetin and iron
Kidney Int 1999; 55: 49-56
31. Kalyani P, Vijaya S and Ramasarma T
Characterization of oxygen free radicals generated during vanadate-
stimulated NADH oxidation
Mol Cell Biochem 1992; 111: 33-40
32. Klahr S
Oxygen radicals and renal diseases
Miner Electrolyte Metab 1997; 23: 140-143
33. Knudsen A, Tappel AL L and North JA
Multiple antioxidants protect against heme protein
and lipid oxidation in kidney tissue
Free Rad Biol Med 1996; 20: 165-173
34. Koksoy FN, Kose H, Soybir GR, Yalcin O, Cokneseli B.
The prohylactic effects of superoxide dismutase, catalase, desferrioxamine,
verapamil and disulfiram in experimental colitis
J R Coll Surg Edinb. 1997; 42: 27-30
35. Kooistra MP, van Es A, Struyvenburg A, Marx JJM
Low iron absorption in erythropoietin-treated hemodialysis patients
J Am Soc Nephrol 1995; 6: 543
Page 61
61
36. Kuhnlein U.
Disulfiram inhibits DNA breakage by hydroxyradical-producing agents
Biochim Biophys Acta 1980; 26: 75-83
37. Lacy F, Gough D and Schmid-Schönbein G
Role of xanthine in hydrogen peroxide production
Free Rad Biol Med 1998; 25: 720-727
38. LeBel CP, Ischiropoulos H, Bondy SC
Evaluation of the probes 2´,7´-dichlorofluorescin as an indicator
of reactive oxygen species formation and oxidative stress
Chem Res Toxicol 1992; 5: 227
39. Leoncini G, Signorello MG, Piana A, Carruba M, Armani U
Hyperactivity and increased hydrogen peroxide formation in
platelets of NIDDM patients
Thromb Res 1997; 86: 153-60
40. Liochev SI and Fridovich I
Superoxide generated by glutathione reductase initiates a vanadate-
dependent free radical chain oxidation of NADH
Arch Biochem Biophys 1992; 294: 403-406
41. Liu D, Liu J and Wen J
Elevation of hydrogen peroxide after spinal cord injury detected
by using the fenton reaction
Free Rad Med 1999; 27: 478-482
42. Loughrey CM, Young IS, Lightbody JH,McMaster D, McNamee PT,
Trimble ER.
Oxidative stress in haemodialysis
QJM 1994; 87: 679-683
Page 62
62
43. Lowe G, Hulley Ch, Rhodes E, Young A and Bilton R
Free radical stimulation of tyrosine kinase and phosphatase activity in
human peripherial blood mononuclear cells
Biochem Biophys Res Commun 1998; 245: 17-22
44. Luciak M, Trznadel K
Free oxygen species metabolism during hemodialysis with different
membranes
Nephrol Dial Transplant Suppl 1991; 3: 66-70
45. Mankhetkorn S, Abedinzadeh Z and Houee-Levin CH
Antioxidant action of sodium diethyldithiocarbamate:
reaction with hydrogen peroxide and superoxide radical
Free Radical Biol Med 1994; 17: 517-527
46. Martin-Mateo MC, Sanchez-Portugal M, Iglesias S, de Paula A and
Bustamante J
Oxidative stress in chronic renal failure
Ren Fail 1999; 21: 155-67
47. Mashiba H and Matsunaga K
Augmented inhibition of tumor cell proliferation in the combined
use of disulfiram with ascorbic acid and the involvement of
oxygen free radicals
Japan J Exp Med 1990; 60: 209-214
48. Matzke GR
Intravenous iron supplementation in end-stage renal disease patients
Am J Kidney Dis 1999; 33: 595-597
49. McCord J
Superoxide dismutase: Rationale for use in reperfusion injury and
inflammation
J Free Rad Biol Med 1986; 2: 307-310
Page 63
63
50. Means RT, Krantz SB
Progress in understanding the pathogenesis of the anemia of chronic disease
Blood 1992; 80: 1639-1647
51. Mira ML, Silva MM, Queiroz MJ, Manso CF
Angiotensin converting enzyme inhibitors as oxygen free radical
scavengers
Free Rad Res Comms 1998; 19: 173-181
52. Monteiro H, Winterbourn Ch and Stern A
Tetravalent vanadium releases ferritin iron which stimulates vanadium-
dependent lipid peroxidation
Free Rad Res Comms 1991; 12-13: 125-129
53. Navarro JF, Teruel JL
Sodium ferric gluconate and iron requirements in hemodialysis patients
Clin Nephrol 1997;48: 202
54. Navarro JF, Teruel JL, Liano F, Marcen R, Ortuno J
Effectiveness of intravenous administration of Fe-gluconate-Na
complex to maintain adequate body iron stores in hemodialysis patients
Am J Nephrol 1996;16: 268-72
55. NKF-DOQI clinical practice guidelines for hemodialysis adequacy
Am J Kidney Dis 1997; 42: 198-202
56. Nissenson AR
Achieving target hematocrit in dialysis patients: new concepts in iron
management
Am J Kidney Dis 1997; 30: 907-11
Page 64
64
57. Nissenson AR, Lindsay RM, Swan S, Seligman P, Strobos J
Sodium ferric gluconate complex in sucrose is safe and effective in
hemodialysis patients: North american clinical trial
Am J Kidney Dis 1999; 33: 471-82
58. Nissenson A and Strobos J
Iron deficiency in patients with renal failure
Kidney Int 1999; 55: 18-21
59. Norlund L, Granvist K, Hansson HA and Norlund R
Diethyldithiocarbamate, but not disulfiram, inhibits alloxan-
induced dye accumulation of isolated mouse islet ß-cells
Med Biol 1986; 64: 37-41
60. Novogrodsky A, Weisspapir M, Patya M, Meshorer A
and Vanichkin A
Tyrphostin 4-nitrobenzylidene malononitrile reduces chemotherapy
toxicity without impairing efficacy
Cancer Res 1998; 58: 2397-2403
61. Orie N. Nelson, Zidek Walter and Tepel Martin
Chemoattractant-and mitogen-induced generation of reactive oxygen species in
human lymphocytes: The role of calcium
Experimental Physiol 1999; 84: 515-520
62. Paller MS and Eaton JW
Hazards of antioxidant combinations containing superoxide dismutase
Free Rad Biol Med 1995; 18: 883-890
63. Pascual J, Teruel JL, Liano F. Sureda A, Ortuno J
Sodium ferric gluconate complex given intravenously for iron
deficiency in hemodialysis
Clin Nephrol 1991; 35: 87
Page 65
65
64. Petrat F, de Groot H, Rauen U
Determination of the chelatable iron pool of single intact cells
by laser scanning microscopy
Arch Biochem Biophys 2000; 376: 74-81
65. Ponka P
Cellular iron metabolism
Kidney Int 1999; 55: 2-11
66. Ratych RE and Bulkley GB
Free-radical-mediated postischemic reperfusion injury in the kidney
J Free Rad Biol Med 1986; 2: 311-319
67. Rehan A, Johnson K, Kunkel R and Wiggins R
Role of oxygen radicals in phorbol myristate acetate-induced
glomerular injury
Kidney Int 1985; 27: 503-511
68. Roob J, Khoschsorur G, Tiran A
Vitamin E attenuates oxidative stress induced by intravenous iron in
patients on hemodialysis
J Am Soc Nephrol 2000; 11: 539-549
69. Schettler V, Wieland E, Methe H, Schuff-Werner P and
Müller GA
Oxidative stress during dialysis: effect on free radical scavenging enzyme
( FRSE ) activities and glutathione ( GSH ) concentration in granulocytes
Nephrol Dial Transplant 1998; 13: 2588-2593
70. Schmidtmann S, Müller M, Baehr R and Precht K
Changes of antioxidative homeostasis in patients on chronic
haemodialysis
Nephrol Dial Transplant Suppl 1991; 3: 71-74
Page 66
66
71. Shah
Role of reactive oxygen metabolites in experimental glomerular disease
Kidney Int 1989; 35: 1093-1106
72. Shi X and Dalal NS
Hydroxyl radical generation in the NADH/microsomal reduction of
vanadate
Free Rad Res Comms 1992; 17: 369-376
73. Silverberg D, Blum M, Agbaria Z, Schwartz D,
Zubkov A., Yachnin T. and Iaina A.
Intravenous iron for the treatment of predialysis anemia
Kidney Int 1999; 55: 79-85
74. Steiner MG and Babbs CH
Hydroxyl radical generation by postischemic rat kidney slices in vitro
Free Rad Biol Medicine 1998; 9: 67-77
75. Sweet Ch, Bradstreet D, Berman R, Jallard N, Weidler D
Pharmacodynamic activity of intravenous E-3174, an angiotensin II,
antagonist in patients with essential hypertension
Am J Hypertension 1994; 7: 1035-1040
76. Tamaki T, Nishiyamaa A, Yoshida H
Effects of EXP3174, a non-peptide angiotensin II receptor antagonist,
on renal hemodynamics and renal function in dogs
Europ J Pharmacol 1993; 23: 15-21
77. Taylor JE, Peat N, Porter C, Morgan AG
Regular low-dose intravenous iron therapy improves response to
erythropoietin in hemodialysis patients
Nephrol Dial Transplant 1996; 1: 1079-1083
Page 67
67
78. Teruel JL, Marcen R, Navarro JF, Villafruela JJ, Fernandez Lucas M
Rivera M, Ortuno J
Influence of body iron stores on the serum erythropoietin concentration in
hemodialyzed patients
Am J Nephrol 1994; 14: 95-98
79. Túri S, Németh I, Torkos A and Sághy L
Oxidative stress and antioxidant defense mechanism in glomerular diseases
Free Rad Biol Med 1997; 22: 161-168
80. Uysal F, Girgin F, Tüzün S, Aldemir S and Yildirim E
Effect of vitamin E on antioxidant enzymes and nitric oxide in ischemia-
reperfused kidney injury
Biochem and Mol Biol Int 1998; 44-47
81. Vijaya S, Crane FL and Ramasarma T
A vanadate-stimulated NADH oxidase in erythrocyte membrane generates
hydrogene peroxide
Mol Cell Biochem 1984; 62: 175-185
82. Vogel S.
Benefits of early utilization of intravenous iron
Nephrol Nurs J 2000; 27: 61-5
83. Walker PD and Shah SV
Hydrogen peroxide cytotoxicity in LLC-PK cells:
A role for iron
Kidney Int 1991; 40: 891-898
84. Watanbe S
In vivo fluorometric measurement of cerebral oxidative stress using
2`-7`-dichlorofluorescein (DCF)
Keio J Med 1998;47: 92-8
Page 68
68
85. Weiss G
Iron and anemia of chronic disease
Kidney Int 1999; 55: 12-17
86. Wolgast M, Bayati A, Hellberg O, Källskog Ö, Nygren K, and Öjteg G
Oxygen radicals in postischaemic damages in the kidney
Klin Wochenschr 1991; 69: 1077-1082
87. Wood MA and Hess MJ
The effects of dihydroxyfumarate on isolated rabbit papillary muscle
function: evidence for an iron dependent non-hydroxyl radical mechanism
Mol Cell Biochem 1987; 78: 161-167
88. Yorgin PD, Belson A, Sarwal M, Alexander SR
Sodium ferric gluconate therapy in renal transplant and renal
failure patients
Pediatr Nephrol 2000; 15: 171-175
89. Yoshioka T, Bills T
Role of intrinsic antioxidant enzymes in renal oxidant injury
Kidney Int 1990; 38: 282-288
90. Zanen AL, Adriaansen HJ, Posthuma R, de Jong TH
`Oversaturation` of transferrin after intravenous ferric gluconate
( Ferrlecit®) in haemodialysis patients
Nephrol Dial Transplant 1996 ;11: 820-824
91. Zhao A, Wu ZQ, Pollack M, Rollwagen FM, Hirszel P, Zhou X
Disulfiram inhibits TNF-alpha-induced cell death
Cytokine. 2000; 12: 1356-67
Page 69
69
9. DANKSAGUNGEN
Herrn Prof. Zidek danke ich für die Ermöglichung der Durchführung dieser
Doktorarbeit.
Mein besonderer Dank gilt Professor Dr.med M.Tepel für die Einarbeitung in die
verschiedenen Arbeitsmethoden, die ausführlichen Diskussionen über aufgetretene
Probleme und Resultate und für sein Engagement bei der Durchführung und bei der
Erstellung dieser Doktorarbeit.
Page 70
70
10. LEBENSLAUF
Angaben zur Person
Name Anna Matuszczyk
Anschrift Steinheimer Str. 12
45896 Gelsenkirchen
Geburtsdatum 10.04.1975.
Geburtsort Ruda/Kattowitz/Polen
Staatsangehörigkeit deutsch
Schulausbildung
1982-1986 Grundschule in Gelsenkirchen
1986-1995 Max Planck-Gymnasium
in Gelsenkirchen-Buer
1995 Abitur
Hochschulausbildung
10/1995 Beginn des Medizinstudiums an
der Ruhr-Universität in Bochum
9/1997 Ärztliche Vorprüfung
8/1998 Erstes Staatsexamen
12/1998-2/2001 Durchführung des
experimentellen Teils der
Doktorarbeit
9/2000 Zweites Staatsexamen
9/2000-9/2001 Praktisches Jahr
in Herne und im Hôpital de
Hautepierre in Straßburg
11/2001 Drittes Staatsexamen
Page 72
This document was created with Win2PDF available at http://www.daneprairie.com.The unregistered version of Win2PDF is for evaluation or non-commercial use only.