Acta histochem. 82, 199-209 (1987) VEB Gustav Fischer Verlag Jena Pathologisches Institut des Bezirkskrankenhauses Garlitz, DDR Vergleichende immunhistochemische und lectinhistochemische Untersuchungen an den Ependymzellen und den Epithelien der Plexus choroidei Comparative immunohistochemical and lectin histochemical study of ependyma and choroid plexus Von MICHAEL KASPER, PETER STOSIEK und ROLAND GOERTCHEN Mit 6 Abbildungen (Eingegangen am 15. August 1986) Summary In a previous investigation we reported on the distribution of intermediate filaments in normal epithelium of choroid plexus and ependyma (KASPER et al. 1986). The present paper describes the results obtained with an enlarged panel of monoclonal antibodies against intermediate fila- ments and additionally with lectins. Ependymal cells contain GF AP and vimentin filaments, whereas epithelial plexus cells express cytokeratin, vimentin and neurofilament 200 KD. Using Concanavalin A, we found a strong cytoplasmatic staining of plexus epithelia and a reduced or failed reaction in ependymal cells. The differences in both cell types may find an explanation in their function or are determined during the ontogenic development. Einfiihrung Vergleichende Studien zur Morphologie von Plexus-Epithelzellen und den Epen- dymzellen der Hirnventrikel existieren schon seit den friihesten Untersuchungen von ASCANAZY (1914) und SCHMID (1929). So vermerkte JAKOB (1952) ausdriicklich histologische Unterschiede zwischen beiden ineinander iibergehenden Epithelien und den von ihnen abgeleiteten Tumoren (z. B. Fehlen von Gliafasern im Plexus-Epithel). Histologische Differenzen zwischen diesen Strukturen fanden auch SHANTA und MANOCHA (1968) bei Ratten und Affen. Nach KRISCH und LEONHARDT (1984) handelt es sich beim Plexus-Epithel urn modifizierte ependymale Zellen bzw. ist das Epithel ependymaler (neuroektodermaler) Herkunft (BARGMANN et al. 1982), wobei die ependymalen Zellen aus 2 grundsatzlichen Zelltypen bestehen: 1. Kinocilienarmen Tanycyten und 2. cuboidalen Ependymzellen mit starkem Kinocilienbesatz (= Ependymocyten). Von anderen Autoren wird auch von Tanycytenependym und Wimpernependym gesprochen (PASSIA et al. 1978). Die Plexus-Epithelzellen haben entweder keine Kino- cilien oder aber zentrale Cilienbiischel und nehmen daher eine Mittelstellung zwischen den o. g. Zelltypen ein (LEONHARDT 1980).
Transcript
Acta histochem. 82, 199-209 (1987)
VEB Gustav Fischer Verlag Jena
Pathologisches Institut des Bezirkskrankenhauses Garlitz, DDR
Vergleichende immunhistochemische und lectinhistochemische Untersuchungen an den Ependymzellen und den Epithelien
der Plexus choroidei
Comparative immunohistochemical and lectin histochemical study
of ependyma and choroid plexus
Von MICHAEL KASPER, PETER STOSIEK und ROLAND GOERTCHEN
Mit 6 Abbildungen
(Eingegangen am 15. August 1986)
Summary In a previous investigation we reported on the distribution of intermediate filaments in normal
epithelium of choroid plexus and ependyma (KASPER et al. 1986). The present paper describes the results obtained with an enlarged panel of monoclonal antibodies against intermediate filaments and additionally with lectins. Ependymal cells contain GF AP and vimentin filaments, whereas epithelial plexus cells express cytokeratin, vimentin and neurofilament 200 KD. Using Concanavalin A, we found a strong cytoplasmatic staining of plexus epithelia and a reduced or failed reaction in ependymal cells. The differences in both cell types may find an explanation in their function or are determined during the ontogenic development.
Einfiihrung
Vergleichende Studien zur Morphologie von Plexus-Epithelzellen und den Ependymzellen der Hirnventrikel existieren schon seit den friihesten Untersuchungen von ASCANAZY (1914) und SCHMID (1929). So vermerkte JAKOB (1952) ausdriicklich histologische Unterschiede zwischen beiden ineinander iibergehenden Epithelien und den von ihnen abgeleiteten Tumoren (z. B. Fehlen von Gliafasern im Plexus-Epithel). Histologische Differenzen zwischen diesen Strukturen fanden auch SHANTA und MANOCHA (1968) bei Ratten und Affen. Nach KRISCH und LEONHARDT (1984) handelt es sich beim Plexus-Epithel urn modifizierte ependymale Zellen bzw. ist das Epithel ependymaler (neuroektodermaler) Herkunft (BARGMANN et al. 1982), wobei die ependymalen Zellen aus 2 grundsatzlichen Zelltypen bestehen:
1. Kinocilienarmen Tanycyten und 2. cuboidalen Ependymzellen mit starkem Kinocilienbesatz (= Ependymocyten). Von anderen Autoren wird auch von Tanycytenependym und Wimpernependym gesprochen (PASSIA et al. 1978). Die Plexus-Epithelzellen haben entweder keine Kinocilien oder aber zentrale Cilienbiischel und nehmen daher eine Mittelstellung zwischen den o. g. Zelltypen ein (LEONHARDT 1980).
200 M. KASPER, P. STOSIEK und R. GOERTCHEN
Ein wesentlicher feinstruktureller Unterschied zwischen Plexus-Epithel- und Ependymzelle ist die Versiegelung mit tight junction8, die nur zwischen den PlexusEpithelzellen realisiert wird (BRIGHTMAN 1961). Eine Ausnahme bilden die Tanycyten der circumventricularen Organe (LEONHARDT 1980).
Wir konnten in eigenen Untersuchungen uber die Verteilung der Intermediarfilamente (IF) in den genannten Zellen ihre unterschiedliche Struktur bekraftigen (KASPER et al. 1986). Die Intermediarfilamente (d = 10 nm) sind neben den Mikrofilamenten und den Mikrotubuli Bestandteil des Cytoskelets der Zelle. Sie ermoglichen die Unterscheidung von 6 Zelltypen (OSBORN 1983). Epitheliale Zellen sind durch Cytokeratine, mesenchymale Zellen durch Vimentin und Muskelzellen durch Desmin charakterisiert. Die meisten Nervenzellen besitzen Neurofilamente und die Gliazellen das saure Gliafaserprotein (GFAP). Einigen Zellen in der fruhen Embryonalentwicklung und bestimmten Zellen im Gehirn fehlen die Intermediarfilamente. Sehr selten finden wir auch gleichzeitige Expression mehrerer Filamenttypen [Ubersicht bei GOWN und GABBIANI (1984)].
Filamente sind auch fur Plexus-Epithel- und Ependymzellen in ultrastrukturellen Untersuchungen beschrieben worden (PICO 1963; BRIGHTMAN und PALAY 1963; BARGMANN und KATRITSIS 1966; MELLER und WAGNER 1968; OKSCHE und VAUPEL V. HARNACK 1969; DOHRMANN und Bucy 1970; MICHAELS und TORNHEIM 1980).
Ziel dieser Arbeit war es, den funktionell noch unklaren morphologischen Differenzen dieser ontogenetisch und topographisch engbenachbarten Zellen nachzugehen. Wir haben dabei die Palette der verwendeten Antikorper erweitern und die Reaktion mit Lectinen hinzunehmen konnen. Lectine sind zuckerbindende Proteine, welche in der Lage sind, Zellen zu agglutinieren oder kohlenhydratreiche Makromolekiile zu prazipitieren (ALROY et al. 1984). Ooncanavalia en8iformi8-Agglutinin (Con A) bindet wie Len8 culinan'8-Agglutinin (LCA) D-Mannosyl- und D-Glucosyl-Zuckerreste. Lectine zeigen unterschiedliche Gewebsreaktionen und konnen daher bei der Klassifikation unterschiedlicher Zell- und Gewebstypen hilfreich sein (SEIFERT et al. 1984).
Material und Methoden Antiseren
Polyklonale Antikorper:
1. Antikorper gegen Rinder-Rei,8nerscher Faden vom Kaninchen (Dr. NAUMANN, Leipzig), 1: 100 verdunnt,
2. Antikorper gegen "tissue polypeptide antigen" (TP A), vom Kaninchen (Prof. BJORKLUND, Uppsala, Schweden), 1: 20 verdunnt,
3. Antikorper gegen N eurofilamentprotein (aIle 3 Subtypen) von der Maus (Prof. PERRY, Cleve· land, USA), 1: 200 verdunnt,
4. Antikorper gegen Keratin vom Kaninchen (Prof. SCHULZ, Berlin), 1: 20 verdunnt.
Monoklonale Antikorper (aIle aus dem Maus-System):
Lectine Con A und LCA, FITC markiert (Institut fUr Biotechnologie, Leipzig) und ConA unmarkiert
(Pharmacia, Schweden). Das Lectin wurde in Vorversuchen ausgetestet und 1: 100 verdunnt eingesetzt. Zur Uberpru
fung der Spezifitat wurde Con A fur 1 h bei 4°C mit 0,2 molJl D( + )-Mannose (Merck, BRD) vorinkubiert (entsprechend der Vorschrift von MEINIEL und MEINIEL [1985]).
Imm unhistochemische Methoden 1. Indirekte Immunperoxidasetechnik unter Verwendung eines peroxidase-markierten Ziege
anti-Kaninchenglobulins (Institut fUr Impfstoffe Dessau, DDR) fUr die polyklonalen Kaninchenseren nach del' Originalmethode von NAKANE (1968).
2. Modifizierte Immunperoxidasetechnik der ursprunglichen Bruckentechnik von MASON et al. (1969) mit einem polyklonalen Ziege-anti-Mausglobulin (SIFIN, Berlin) und einem 2. monoklonalen Anti-Peroxidase-Antikorper BL-POD II (Sektion Biowissenschaften der Karl-MarxUniversitat, Leipzig) zur Testung der monoklonalen Antikorper bzw. des polyklonalen MausSerums. Ais Kontrollen dienten irrelevante Hybridomuberstande und phosphatgepufferte Kochsalz16sung (PBS), pH = 7,2.
3. Direkte Fluorescenz mit markiertem Lectin: Inkubation bei Raumtemperatur, 45 min, \Vaschen und Eindecken in Glycerin-PBS unter Zusatz von p-Phenylendiamin (HUFF et al. 1982). Beobachtung unter dem Auflicht (Jenalumar contrast, VEB Carl Zeiss .JENA), HBO 202.
4. Uberschichtung der Schnitte mit Con A in der Konzentration von 0,05 mgJml, 30 min, Raumtemperatur, Waschen (3 X 10 min) und Inkubation mit 0,02 % Meerettichperoxidase (Arzneimittelwerk Dresden). Verwendung von Kontrollen (PBS, Ulex-europaeus-Agglutinin I) und entsprechenden Hemmtests (s. 0.) mit dem korrespondierenden Zucker.
Gewebe Gehirn von Maus, Ratte, Meerschwein und menschliches Plexus- und Ependymgewebe (9 X
Autopsiematerial, 3 bis 6 h post mortem) wurden in Einbettmedium (Jung, BRD) aufgenommen, in flussigern Stickstoff gefroren und davon etwa 4 [tm dicke Kryostatschnitte angefertigt. FUr den Nachweis der Verteilung der IF wurde ein Gehirn (humanes Material) vollstandig zur Praparation allel' Plexus chol'oidei aufgeal'beitet.
Ergebnisse
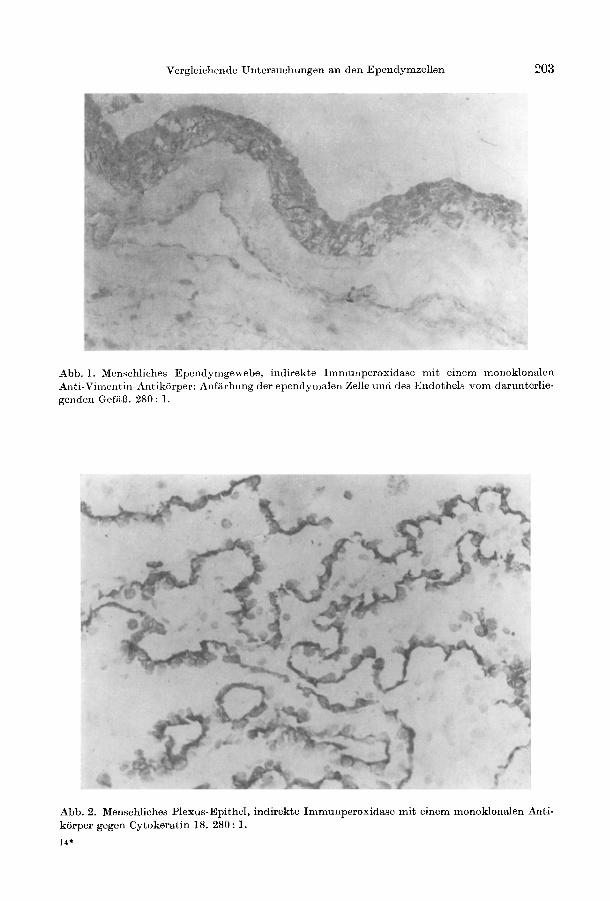
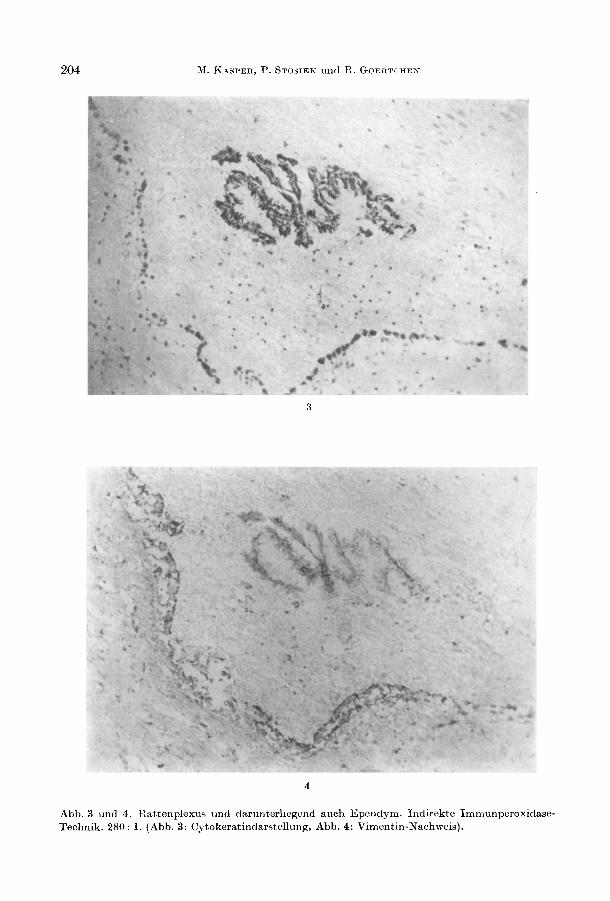
Wahrend sich in den Epithelien des menschlichen Plexus choroideus 3 verschiedene IF - Cytokeratin, Neurofilament und Vimentin - fanden, lieEen sich in den Ependymzellen nur GFAP und Vimentin nachweisen (Tabelle 1, Abb. 1). TPA als Abbauprodukt der Cytokeratine wurde ebenfalls nur im Plexus-Epithel immunhistochemisch lokalisiert. Bei den individuellen Cytokeratinen (MOLL et al. 1982) war die Nr. 19 immer negativ im Gegensatz zur positiven Reaktion des Antikorpers gegen Cytokeratin 18 (Abb.2, Tabelle 1). Die Cytokeratinantikorper markierten auch Maus-, Ratten- und Meerschweinchenplexus (Abb. 3). Bei der Ratte lieE sich aber kein Vimentin im Epithel des Plexus choroideus nachweisen. Lediglich das Bindegewebe unter dem Epithel reagierte positiv (Abb. 4). 1m Ependym der Ratte war Vimentin und fokal Cytokeratin exprimiert (Abb. 3, 4). Die Dreifachexpression von IF lieE sich auch beim menschlichen Plexus fUr alle topographischen Lokalisationen zeigen (Tabelle 2). 14 Acta histochem., Bd. 82
202 1\1:. KASPER, P. STOSIER und R. GOERTCHE::-<
Tabelle 1. Vergleich der Ergebnisse immunhistochemischer Testungen verschiedener Antikorper an menschlichen Geweben
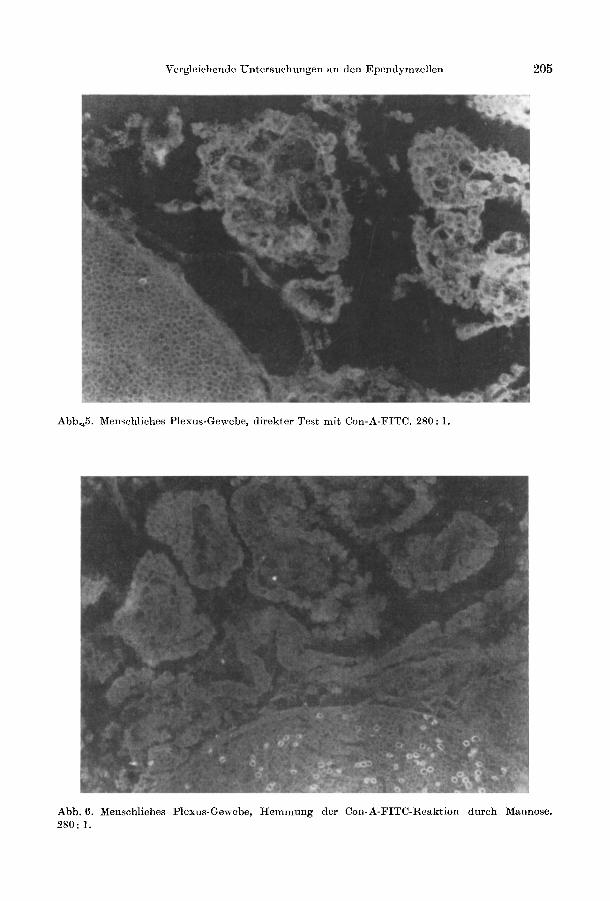
Eine Con A-Bindung durch Glykoproteine der Plexus-Epithelzellen war immer deutlich erkennbar (Abb. 5, Tabelle 3). Die Reaktion umfaEte diffus das Cytoplasma und lieE sich im Hemmtest mit dem korrespondierenden Zucker vollstiindig blockieren (Abb. 6).
In den Ependymzellen war die Reaktion entweder ganz negativ oder geringgradig subependymal vorhanden. Nur in einem Fall zeigten die ependymalen Zellen eine stiirkere Con A-Affinitiit.
Ulex europaeu8-Agglutinin I wurde lediglich an stromalen GefiiEendothelien gebunden. LCA konnte ebenfalls im Plexus-Epithel nachgewiesen werden, wobei das Fluorescenzbild den Con A-Testungen entsprach.
CEA und verwandte Blutgruppenantigene waren in den getesteten Geweben nicht vorhanden. Ebenfalls negativ fiel die Reaktion des Antiserums gegen den Reifinerschen Faden aus.
Diskussion
Die Coexpression von mehreren Intermediarfilamenttypen in einer Zelle wird fUr Zellen in der Kultur (Ubersicht bei OSBORN and' WEBER [1983]), Tumorzellen (HOLTHOFER et al. 1983, KREPLER et al. 1982, STOSIER und GOERTCHEN 1986), aber auch fUr Normalgewebe (HOLTHOFER et al. 1984, LANE et al. 1983, KASPER et al. 1986) beschrieben.
Sie ist insgesamt selten und scheint dort vorzukommen, wo differente Gewebsprogramme ortlich oder ontogenetisch eng benachbart sind oder Gewebe bzw. Tumoren eine ihrem Keimblattprogramm differente Differenzierung erfahren. Unklar ist, ob es sich dabei urn ein echtes Nebeneinander aller Filamente handelt oder ob Heteropolymere aus verschiedenen Filamenttypen gebildet werden konnen (STEINERT et al. 1985). Die Tatsache, daB im Plexusepithel nur ein Subtyp der
Vergleichende Untersuchungen an d en Ependymzellen :!03
Abb. 1. Menschliches Ependymgewebc, indirekte Immunperoxidase mit einem monoklonalen Anti-Vimentin- Antikiirper: Anfiirbung der ependymalen Zelle und des Endothels yom darunterliegenden Gefiif3. 280: 1.
Abb. 2. Menschliches Plexus-Epithel, indirekte Immunperoxidase mit einem monoklonalen Antikiirper gegen Cytokeratin 18. 280: 1.
]4*
204 l\I. KASPER, P. STO,IEK uncI It. GOEUTCHEN
3
4
Abb. 3 und 4. Rattenplexus und darunterliegend auch Ependym. Indirekte ImmunperoxidaseTechnik. 280: 1. (Abb. 3: Cytokeratindarstellung, Abb. 4: Vimentin·Nachweis).
Vergleichende Untersuchungen an den Ependymzellen 205
Abb.4 5. Menschliches Plexus-Gewebe, direkter Test mit Con-A-FITC. 280: 1.
Abb.6. Menschliches Plexus-Gewebe, Hemmung der Con-A-FITC-Reaktion durch Mannose. 280: 1.
206 JVL KASPER, P. STOSIEK und R. GOERTCHEN
Tabelle 3. Positiver Nachweis von Con A.Bindung
Immunfluorescenz Peroxidasetechnik
Plexusepithel
Ependym
9/9
0/4
5/5
1/3
Tabelle 4. Zusammenfassende Darstellung der Differenzen zwischen Ependymzellen und PlexusEpithelzellen im menschlichen Gehirn
IF
• Cytokeratin
• GFAP
• Neurofilament 200KD
TPA
Con A·Bindung
Praealbumin
S·100 Protein
Plexus.Epithel
+ + ,
T
+
+ + +
Ependymzelle
KASPER et al. (1986) SCHWECHHEIMER et al. (1985) MIETTINEN et al. (1986)
+/- ARilWND et al. (1980)
+ GOWN und GABBIANI (1984)
NAll'HRATH et al. (1985)
-~/-
ALESHIRE et al. (1983)
+ YAMAGUCHI (1980)
sonst nur als Triplet vorkommenden Neurofilamente in der Coexpression vorkommt (mit mehreren monoklonalen Antikorpern von uns gesichert), spricht eher fur die Annahme der Heteropolymere.
Unsere Untersuchungen zur Verteilung der Filamente in den Plexus-Epithel· und Ependymzellen zeigen eine mehr "neuroepitheliale" Differenzierung der Plexus·Epithelien (Cytokeratin und Neurofilament) und eine mehr "makrogliose" Differenzierung (GFAP und Vimentin) der Ependymzellen (Tabelle 1).
Moglicherweise wird die Expression der Filamente in einer Zelle, die in der Keimbahn in einem common ancestral gene angelegt ist (KRIEG et al. 1985) in den fruhen Stadien der Ontogenese durch andere Zellen induziert. Hier konnte die subependymale Gewebeplatte eine Rolle spielen. Bei den choroidalen Organen (VIGH 1971) ist es die leptomeningeale Unterlage, beim Ependym der Ventrikelauskleidungen im wesentlichen die 1- oder 2schichtige Glia. Vollstandig offen ist aber immer noch, ob ausschliel.llich die differentielle Genaktivitiit einer "ependymalen Stammzelle" oder aber funktionelle Aspekte im Vordergrund stehen.
Regionale Unterschiede in der Expression der IF zwischen Plexus der Seitenventrikel, des III. und des IV. Ventrikels bestehen nicht (Tabelle 2).
Die Angaben zur Coexpre~sion von GFAP und Vimentin im Ependym stimmen mit Untersuchungen anderer Autoren uberein (DE ARMOND et al. 1980, SHAW et al. 1981, GOWN und GAB· BIANI 1984).
Die Dreifachexpression wurde unseres Wissens noch nicht beschrieben. 1m Plexusepithel lal.lt sich GFAP nicht nachweisen. Nur in einigen Plexus·Papillomen ist GFAP fokal zu finden (ENG und RUBINSTEIN 1978; RUBINSTEIN und BRUCHER 1981). Interessanterweise ist im Plexus von Neugeborenen bei der sogenannten "Ependymzotte" (BECKER 1939) isoliert GF AP nachweisbar (eigene unveroffentlichte Untersuchungen). Unklar ist die geringe Cytokeratinexpression im Ependym der Ratte (Abb. 3), die auch von MIE'l''l'INEN et al. (1986) fUr Maus· und Rattenependym nach-
Vergleichende Untersuchungen an den Ependymzellen 207
gewiesen wurde. Von SCHWECHHEIMER et al. (1985) und MIETTINEN et al. (1986) sind biochemisch die individuellen Cytokeratine 8, 18 und 19 im Plexus-Epithel erfal.!t worden. Mit 3 monoklonalen Anti-Cytokeratin 19-Antikorpern haben wir in keinem FaIle eine positive immunhistochemische Reaktion erhalten.
In Tabelle 4 sind zusammenfassend wesentliche Unterschiede zwischen Plexus-Epithel und Ependym dargestellt. Daneben gibt es noch eine Reihe anderer Differenzen in tierischen Geweben, z. B. die Anwesenheit von Heymann-Antigen in ependymalen Zellen der Ratte (Plexus.Epithel negativ, ASSMAN et al. [1986]) oder die unterschiedliche Anfarbbarkeit der Gewebe mit einer modifizierten Bodian-Versilberung von GAMBETTI et al. (1981), eigene unveroffentlichte Ergebnisse.
Die Untersuchungen von MEINIEL und MEINIEL (1985) uber die,Bindung von Con A an das Subcommissuralorgan von Vertebraten (SCO), eine auf Sekretion des Rei/3nerschen Fadens spezialisierte ependymale Zellpopulation (STERBA et al. 1981), gab Anlal.! zur Prufung der Verhaltnisse im menschlichen Plexus, da dort im Vergleich zum ubrigen Ependym erhohte sekretorische Aktivitaten nachweisbar sind (OKSCHE 1984) und im adulten Gehirn beim Menschen kein SCO vorhanden ist. Eine Con A-Bindung ist bisher nur von WARCHOL (1978) auf ultrastrukturellem Niveau am apical en Saum der Ependymzellen gezeigt worden. Die eigenen Befunde (Abb. 5) ergaben eine starke cytoplasmatische Con A-Bindung im Plexus-Epithel (neben anderen ausgewiesenen Con AReceptor tragenden Strukturen). Die Ergebnisse am Plexus - dem wesentlichen Liquorproduzenten des Gehirns - korrespondieren mit der bevorzugten Con A-Bindung des eben falls sekretorisch aktiven SCO.
Das Vorkommen derartiger Kohlenhydratstrukturen als Bestandteil von cytoplasmatischen Glykoproteinen konnte fUr Produktion und Regulation des Liquors von Bedeutung sein (BATENKO et al. 1984). Weitere Lectinbindungsstudien sind von MARES et al. (1984) mit Weizenkeim-Agglutinin (Plexus und Ependym positiv) und mit Erdnul.llectin (VIERBUCHEN und KLEIN 1983) durchgefUhrt worden. Der positive Nachweis des letzteren im Plexus-Epithel ist an Neuraminidase behandeltes Gewebe gebunden. Die Testungeines Antiserums gegen den Rei/3nerschen Faden ergab negative Resultate in den eigenen Untersuchungen. Immunreaktives Material findet sich jedoch in den Plexus-Epithelzellen vom Huhn (RODRIGUEZ et al. 1984).
Zusammenjassend kann festgestellt werden, dal.! sich die von uns bereits auf der Ebene der IF beschriebenen Differenzen zwischen Plexus-Epithel und Ependym (KASPER et al. 1986) bestatigen und auf die Ebene der Lectinbindungen ausweiten liel.!en. Welche morphologische funktionelle und ontogenetische Bedeutung diese interessanten Befunde haben, ist noch vollig offen und mul.! Gegenstand weiterer Untersuchungen sein.
Danksagung: Fur die freundlicherweise bereitgestellten poly- und rnonoklonalen Antikorper bedanken wir uns bei allen genannten Kollegen. Fur die gute technische Assistenz danken wir Frau M. HAASE und Frau B. LECHNER.
Literatur
ALESHIRE, S. L., BRADLEY, C. A., RICHARDSON, L. D., and PARL, F. F., Localization of human prealbumin in choroid plexus epithelium. J. Histochem. Cytochem. 31,608-612 (1983).
ALROY, .r., UCCI, A. A., and PEREIRA, M. A. E., Lectins: histochemical probes for specific carbohydrate residues. In: Advances in Immunhistochemistry (R. A. DE LELLIS, ed.). Masson, New York 1984, pp. 67-68.
ASOANAZY, M., Zur Physiologie und Pathologie der Plexus chorioidei. Zbl. Path. 25, Erg. H. 85 (1914).
ASSMANN, K. J. M., LANGE, W. P. H., TANGELDER, M. M., and KRONE, R. A. P., The organ distribution of gp-330 (Heymann antigen) and gp-90 of the mouse and the rat. Virchows Arch. (Pathol. Anat.) 408, 541-553 (1986).
BARGMANN, W., und KATRITSIS, E., tiber die sogenannten Filamente und das Pigment im Plexus choroideus des Menschen. Z. Zellforsch. 75, 366-370 (1966).
- OSCHKE, A., FIX, J. D., and HAYMAKER, W., Meninges, choroid plexus, ependyma and their reactions. Part 1. Histology and functional considerations, pp. 560-641. In: W. HAYMAKER, R. D. AMADS (eds.), Histology and Histopathology of the Nervous System. Vol. 1. Springfield, Ill.: Ch. C. Thomas 1982.
208 M. KASPER, P. STOSIEK und R. GOERTCHEN
BA'i'ENKO, L.1., IVASHERSKAIA, E. B., and PEREKHVALSKAIA, T. V., Histochemical analysis of the glucosaminoglucan content in the choroid plexus during hydration and dehydration. Bjull. eksp. BioI. Med. 97, 504-505 (1985).
BEcKER, G., Beitriige zur Orthologie und Pathologie der Plexus chorioidei und des Ependyms. Beitr. path. Anat. 103,457-478 (1939).
BRIGH'i'MAN, M. W., The fine structure of ciliated ependyma. Anat. Rec. 139, 210-211 (1961). - and PALAY, S. L., The fine structure of ependyma in the brain of the rat. J. Cell BioI. 19, 415
to 439 (1963). DE ARMOND, F., ENG, S. J., and RUBINS'i'EIN, L. J., The application of glial fibrillary acidic (GFA)
protein immunohistochemistry in neurooncology. Path. Res. Pract. 168,374-394 (1980). DOHRMANN, G. J., and Bucy, P. C., Human choroid plexus: a light and electron microscopic study.
J. Neurosurg. 33,506-516 (1970). ENG, L. F., and RUBINSTEIN, L. J., Contribution of immunohistochemistry to diagnostic problems
of human cerebral tumors. J. Histochem. Cytochem. 26, 513-522 (1978). GAMBE'i''I'I, P., AU'i'ILIO·GAMBE'i''i'I, L., and PAPASOZOMENOS, S. C., Bodian's silver method stains
neurofilament polypeptides. Sciences 213, 1521-1522 (1981). GOWN, A. M., and GABBIANI, G., Intermediate-sized (10 nm) filaments in human tumors. In:
DE LELLIS, R. A. (ed.): Advances in Immunohistochemistry. Masson, New York 1984, pp. 89-109.
HOL'i'HOFER, H., MIET'i'INEN, A., LEH'i'O, V.-P., LEH'i'ONEN, E., and VIR'i'ANEN, J., Expression of vimentin and cytokeratin types of intermediate filament proteins in developing and adult human kidneys. Lab. Invest. 50, 552-559 (1984). - PAASIVUO, R., LEH'i'O, U.-P., LINDNER, E., ALF'i'HAN, 0., and VIR'i'ANEN, J., Cellular origin and differentiation of renal carcinomas: A fluorescence microscopic study with kidneyspecific antibodies, antiintermediate filament antibodies, and lectins. Lab. Invest. 49, 317 to 326 (1983).
HUFF, J. C., WES'i'ON, W. L., and WANDA, K. D., Enhancement of specific immunofluorescent findings with use of a paraphenylenediamine mounting buffer. J. Invest. Dermatol. 78, 449 to 450 (1982).
JAKOB, H., Gut- und bosartige Geschwulste des Plexus chorioideus. Arch. Psychiatr. Neurol. 189, 162-180 (1952).
KASPER, M., STOSIEK, P., and KARS'i'EN, U., Detection of cytokeratin(s) in epithelium of human Plexus chorioideus by monoclonal antibodies. Acta histochem. 78, 101-103 (1986).
KREPLER, R., DENK, H., AR'i'LIEB, U., and MOLL, R., Immunocytochemistry of intermediate filament proteins present in pleomorphic adenomas of the human parotid gland: Characterization of different cell types in the same tumor. Differentiation 21, 191-198 (1982).
KRIEG, T. M., SCHAFER, M. P., CHENG, C. K., FILPULA, D., FLAHER'i'Y, P., S'i'EINER'i', P. M., and Roop, D. R., Organization of a type I keratin gene: Evidence for evolution of intermediate filaments from a common ancestral gene. J. BioI. Chem. 260, 5867-5879 (1985).
LEONHARDT, H., Ependym und Circumventriculiire Organe. In: OSCHKE, A., und L. VOLLRA'i'H (Eds.): Handbuch der Mikroskopischen Anatomie des Menschen, Bd.4/10. Springer-Verlag, Berlin-Heidelberg-New York 1980, pp. 177-666.
MARES, V., BORGES, L. F., and SIDMAN, R. L., Uptake and transport of lectins from the cerebrospinal fluid by the immature mouse brain. Acta histochem. 74, 11-19 (1984).
MASON, T. E., PHIFER, R. F., SPICER, S. S., SWALLOW, R. A., and DRESKIN, R. B., An immunoglobulin-enzyme bridge method for localizing tissue antigens. J. Histochem. Cytochem. 17, 563-569 (1969).
MEINIEL, R., and MEINIEL, A., Analysis of the secretions of the subcommissural organs of several vertebrate species by use of fluorescent lectins. Cell Tissue Res. 239, 359-364 (1985).
MICHAELS, J. E., and TORNHEIM, P. A., Fine structure of the lateral areas of the rhombencephalic tela of the bullfrog, Rana catesbeiana. Cell Tissue Res. 211, 449-463 (1980).
MIE'i''i'INEN, M., CLARK, R., and VIR'i'ANEN, 1., Intermediate filament proteins in choroid plexus and ependyma and their tumors. Amer. J. Pat hoI. 123, 213-240 (1986).
Vergleichende Untersuchungen an den Ependymzellen 209
MOLL, R., FRANKE, W. W., SCHILLER, D. L., GEIGER, B., and KREPLER, R., The catalog of human cytokeratins: Patterns of expression in normal epithelia, tumors and cultured cells. Cell 31, 11-24 (1982).
NAKANE, P. K., Simultaneous localization of multiple tissue antigens using the peroxidase-labeled antibody method. A study on pituitary glands of the rat .• 1. Histochem. Ctyochem. 16,557-560 (1968).
NATHRATH, W. B .• T., HEIDENKUMMER, P., BJORKLUND, V., and BJORKLUND, B., Distribution of tissue polypeptide antigen (TP A) in normal human tissues. ,T. Histochem. Cytochem. 33, 99-109 (1985).
OKSCHE, A., Das Plexusepithel: Struktur und Funktion. Verh. Anat. Ges. 78, 71-83 (1984). und VAUPEL V. HARNACK, M., Elektronenmikroskopische Studien tiber Altersveranderungen (Filamente) der Plexus choroidei des Menschen (Biopsiematerial). Z. Zellforsch. 93, 1-29 (1969).
OSBORN, M., Intermediate filaments as histologic markers: An overview. J. Invest. DermatoL 81, 104-107 (1983).
- and WEBER, K., Tumor diagnosis by intermediate filament typing: A novel tool for surgical pathology. Lab. Invest. 48, 372-394 (1983).
PASSIA, D., GOSLAR, H. G., und BITSCH, J., Das enzymhistochemische Verhalten des Tanycytenependyms in III. Ventrikel der Ratte bei Thiaminmangelernahrung. Acta histochem. 61, 72-83 (1978).
PICO, J. E., Die Feinstruktur versenkter Ependymzellen innerhalb von gliasen Narbenbereichen. Acta neuropath. (Berl.) 3, 137-143 (1963).
RODRIGUEZ, E. M., OKSCHE, A., HEIN, S., RODRIGUEZ, S., and YULIS, R., Comparative immunocytochemical study of the subcommissural organ. Cell Tissue Res. 237, 427-441 (1984).
RUBINSTEIN, L. J., and BRUCHER, J.-M., Focal ependymal differentiation in choroid plexus papillomas. An immunperoxidase study. Acta NeuropathoL (Berl.) 53, 29-33 (1981).
SCHMID, H., Anatomischer Bau und Entwicklung der Plexus chorioidei in der Wirbeltierreihe und beim Menschen. Z. mikrosk.-anat. Forsch. II, 413-420 (1929).
SCHWECHHEIMER, K., ACHTSTATTER, T. H., und FRANKE, W. W., Primare epitheliale intracranielle Tumoren. Verh. Dtsch. Ges. PathoL 69, 645 (1985).
SEIFERT, G., DENK, H., KLEIN, P. J., STEIN, H., und OTTO, H. F., Die Anwendung der Immunzytochemie in der praktischen Diagnostik des Pathologen. Pathologe 5, 187-199 (1984).
SHANTA, T. R., and MANOCHA, S. L., Enzymhistochemistry of the choroid plexus in rat and squirrel monkey. Histochemie 14, 149-160 (1968).
SHAW, G., OSBORN, M., and WEBER, K., An immunofluorescence microscopical study of the neurofilament triplet proteins, vimentin and glial fibrillary acidic protein within the adult rat brain. Europ. J. Cell BioI. 26, 68-82 (1981).
STEINER, P. M., STEVEN, A. C., and Roop, D. R., The molecular biology of intermediate filaments. Cell 42, 411-419 (1985).
STERBA, G., KLEIM, J., NAUMANN, W., and PETTER, H., Immunocytochemical investigation of the subcommisural organ in the rat. Cell Tissue Res. 218, 659-662 (1981).
STOSIEK, P., und GOERTCHEN, R., Immunpathologischer Beitrag zum malignen diffusen Mesotheliom. ZbL allg. Pathol. pathoL Anat. 132, 119-128 (1986).
VIERBUCHEN, M., and KLEIN, P. J., Histochemical demonstration of neuraminidase effects in pneumococcal meningitis. Lab. Invest. 48, 181-186 (1983).
VIGH, B., Das Paraventrikularorgan und das zirkumventrikulare System. Studia BioI. Hungar. 10. Budapest: Akademiai Kiad6 1971.
WARCHOL, J. B., Concanavalin A binding sites on the luminal surface of ependymal cells of third ventricle. Histochemistry 58, 139-143 (1978).
YAMAGUCHI, Y., Studies on immunohistochemical localization of S-100 and GFA proteins in the rat nervous system and in human brain tumors. Brain Res. 32, 1055-1064 (1980).
Anschrift der Verfasser: Dr. MICHAEL KASPER, Pathologisches Institut, Bezirkskrankenhaus Garlitz, Girbigsdorfer StraJ3e 1-3, Garlitz, DDR - 8900.