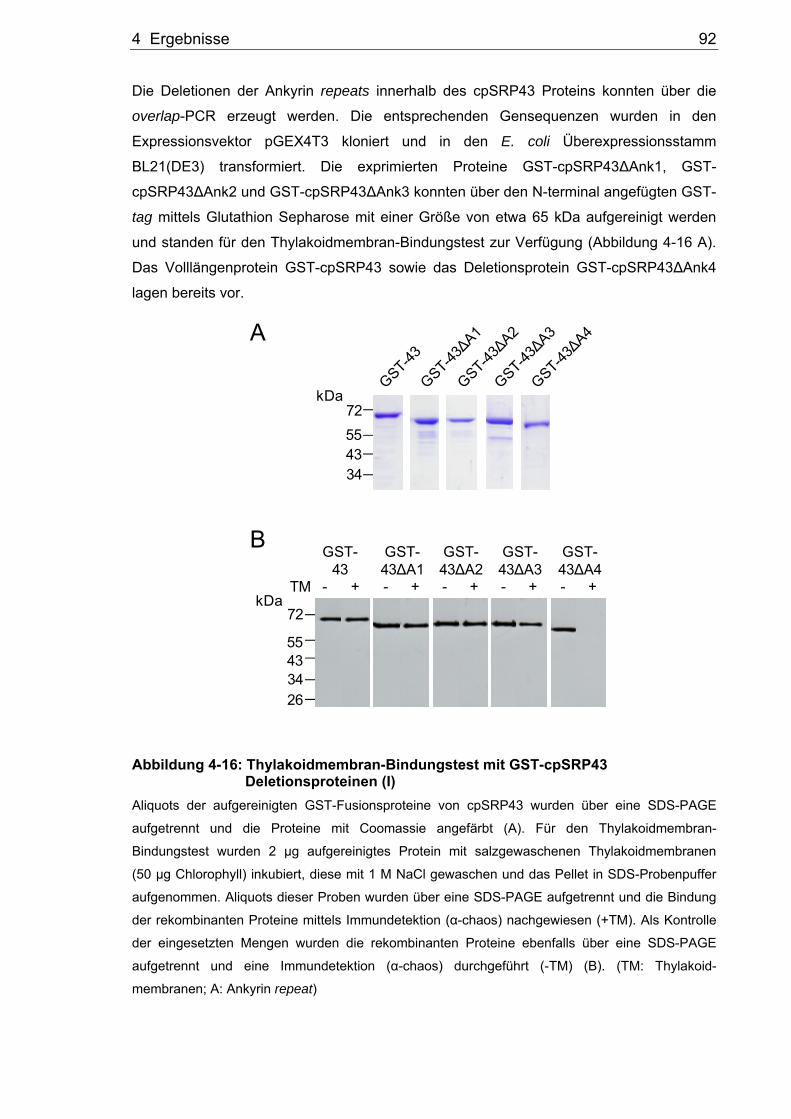
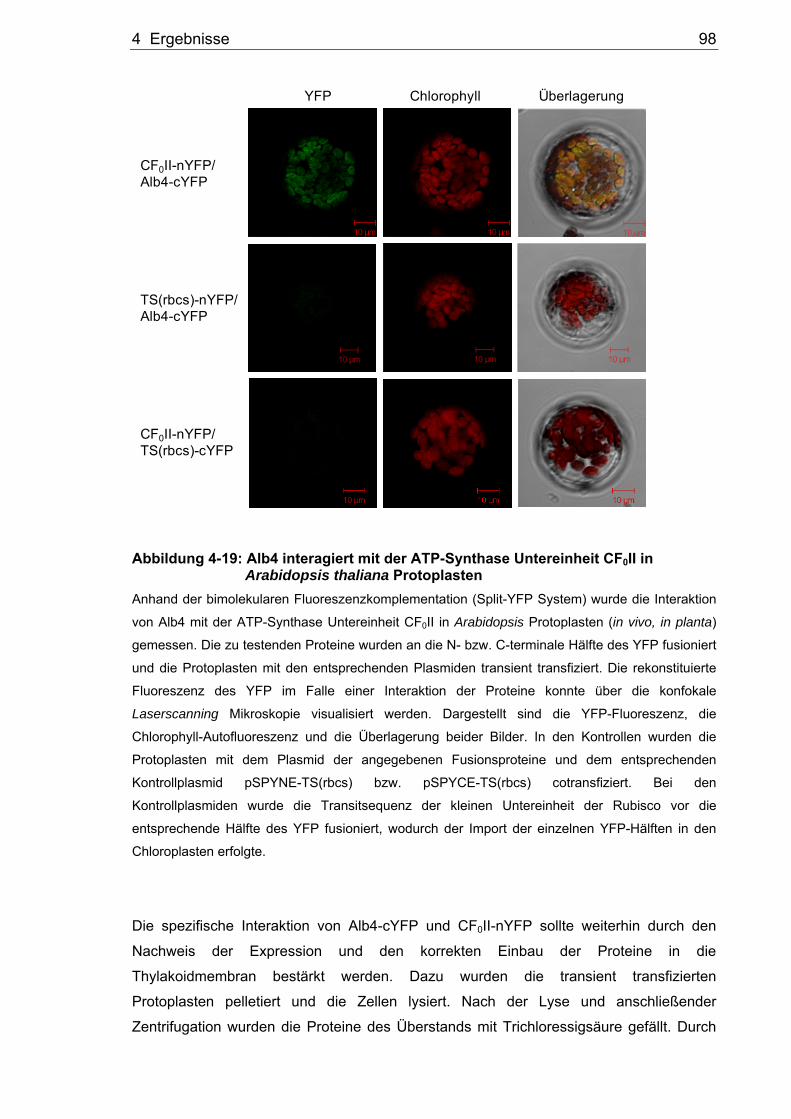
Untersuchungen zur Rolle von Alb3 und Alb4 bei der Biogenese der Thylakoidmembran in Arabidopsis thaliana Dissertation zur Erlangung des Grades eines Doktors der Naturwissenschaften der Fakultät für Biologie und Biotechnologie an der Internationalen Graduiertenschule Biowissenschaften der Ruhr-Universität Bochum angefertigt in der AG Molekularbiologie pflanzlicher Organellen vorgelegt von Thomas Bals aus Unna Bochum Oktober, 2009 Erstgutachterin: Frau Prof. Dr. Danja Schünemann Zweitgutachter: Herr Prof. Dr. Matthias Rögner
Transcript
Untersuchungen zur Rolle von Alb3 und Alb4 bei der Biogenese der Thylakoidmembran
in Arabidopsis thaliana
Dissertation zur Erlangung des Grades
eines Doktors der Naturwissenschaften
der Fakultät für Biologie und Biotechnologie
an der Internationalen Graduiertenschule Biowissenschaften
der Ruhr-Universität Bochum
angefertigt in der
AG Molekularbiologie pflanzlicher Organellen
vorgelegt von
Thomas Bals aus
Unna
Bochum
Oktober, 2009
Erstgutachterin: Frau Prof. Dr. Danja Schünemann
Zweitgutachter: Herr Prof. Dr. Matthias Rögner
Analyses of Alb3 and Alb4 in thylakoid membrane biogenesis
in Arabidopsis thaliana
Dissertation to obtain the degree
Doctor Rerum Naturalium (Dr. rer. nat.)
at the Faculty of Biology and Biotechnology
Ruhr-University Bochum
International Graduate School of Biosciences Ruhr-University Bochum
1.1 Funktion und evolutionärer Ursprung der Chloroplasten.................................1 1.2 Proteintransport kernkodierter Proteine über die chloroplastidären
Hüllmembranen...............................................................................................1 1.3 Proteintransport zu den Thylakoiden ..............................................................2 1.3.1 Proteinimport in das Thylakoidlumen ..............................................................3 1.3.1.1 Der cpSec-Weg...............................................................................................4 1.3.1.2 Der cpTat-Weg................................................................................................4 1.3.2 Proteinintegration in die Thylakoidmembran...................................................5 1.3.2.1 Spontane Insertion ..........................................................................................5 1.3.2.2 Der cpSRP-Weg..............................................................................................5 1.4 Proteininteraktionen der Komponenten des posttranslationalen cpSRP-
Transportwegs ................................................................................................8 1.5 Die Oxa1/YidC/Alb3-Proteinfamilie ...............................................................10 1.5.1 Oxa1..............................................................................................................11 1.5.2 Yid C .............................................................................................................13 1.5.3 Alb3 ...............................................................................................................14
3.6.11 Restriktionsanalyse und Sequenzierung.......................................................37 3.6.12 Agarosegelelektrophorese zur Trennung von DNA.......................................37 3.6.13 Split-Ubiquitin System ...................................................................................38 3.6.13.1 Transformation von Saccharomyces cerevisiae mit Plasmid-DNA ...............39 3.6.13.2 β-Galaktosidase Filtertest .............................................................................39 3.6.14 Bimolekulare Fluoreszenzkomplementation (Split-YFP System) ..................40 3.6.14.1 Isolierung von Arabidopsis thaliana Protoplasten und transiente
Transfektion ..................................................................................................41 3.7 Biochemische Methoden...............................................................................42 3.7.1 Überexpression von Proteinen in Escherichia coli ........................................42 3.7.2 Denaturierende Aufreinigung von Proteinen mit His-tag über Ni-NTA..........42 3.7.3 Native Aufreinigung von Proteinen mit His-tag über Ni-NTA.........................43 3.7.4 Aufreinigung von Proteinen mit GST-tag über Glutathion-Sepharose ..........44 3.7.5 Abspaltung des GST-tags über Thrombinverdau..........................................45 3.7.6 Dialyse von Proteinen ...................................................................................45 3.7.7 BCA-Proteinbestimmung...............................................................................45 3.7.8 SDS-Polyacrylamid-Gelelektrophorese (SDS-PAGE)...................................45 3.7.9 Coomassie-Färbung......................................................................................47 3.7.10 Elektrophoretischer Transfer von Proteinen auf
Nitrocellulosemembranen (Western-Blot) .....................................................47 3.7.11 Proteinfärbung mit Ponceau S ......................................................................47 3.7.12 Immundetektion von Proteinen .....................................................................48 3.7.13 Affinitätsaufreinigung von Antikörpern ..........................................................49 3.7.14 In vitro Transkription......................................................................................50 3.7.15 In vitro Translation.........................................................................................50 3.7.16 In vitro Expression mit RTS 100 Wheat Germ ..............................................51 3.7.17 Pull down Experiment ...................................................................................51 3.7.18 Peptide-Scanning ..........................................................................................52 3.7.18.1 Bindung des rekombinanten Proteins an die PepSpot-Membran .................52 3.7.18.2 Elektrophoretischer Transfer eines rekombinanten Proteins von der
PepSpot-Membran auf eine PVDF-Membran ...............................................53 3.7.18.3 Regeneration der PepSpot-Membran ...........................................................54 3.7.19 Präparation von Thylakoidmembranen aus Arabidopsis thaliana .................55 3.7.19.1 Chlorophyllbestimmung.................................................................................56 3.7.19.2 Salzwaschung der Thylakoidmembranen .....................................................56 3.7.19.3 Solubilisierung der Thylakoidmembranen .....................................................56 3.7.20 Crosslinking der Thylakoidmembranproteine von Arabidopsis thaliana........57 3.7.21 Coimmunpräzipitation ...................................................................................57 3.7.21.1 Kopplung des Antikörpers an Protein-A-Sepharose beads...........................57 3.7.21.2 Präzipitation ..................................................................................................58 3.7.22 Thylakoidmembran-Bindungstest..................................................................58 3.7.23 Thylakoidmembran-Bindungstest mit Peptidinhibierung ...............................59 3.7.24 Expressionsnachweis der YFP-Fusionsproteine in Arabidopsis thaliana
4.1 Herstellung spezifischer Antikörper gegen Alb3 und Alb4 ............................63 4.1.1 Expression der Antigene GST-nAlb3, GST-cAlb3 und cAlb4-his ..................64 4.1.2 Test der Antikörper α-Alb3 und α-Alb4 gegen die Antigene..........................65 4.1.3 Test der Antikörper α-Alb3 und α-Alb4 gegen die in vitro translatierten
Volllängenproteine ........................................................................................67 4.1.4 Test der Antikörper α-Alb3 und α-Alb4 mit Thylakoidmembran-proteinen ....68 4.2 Analyse der Interaktionen von Alb3 und Alb4 mit den Proteinen des
cpSRP-Transportwegs ..................................................................................69 4.2.1 Interaktionen von Alb3 und Alb4 mit den Proteinen des cpSRP-
Transportwegs im Hefe Split-Ubiquitin System.............................................69 4.2.2 Proteininteraktionen von Alb3 und Alb4 mit den Proteinen des cpSRP-
Transportwegs im Split-YFP System ............................................................71 4.2.3 Analyse der Binderegion zwischen Alb3 und cpSRP43 über das Split-
YFP System ..................................................................................................76 4.2.4 Peptide Scanning von Alb3 mit rekombinantem cpSRP43 ...........................80 4.2.5 Bestätigung der mittels Peptide Scanning gezeigten Binderegionen im
Split-YFP System ..........................................................................................82 4.2.6 Verifizierung der Binderegionen im Hefe Split-Ubiquitin System ..................84 4.2.7 Pull down Analysen zur Interaktion von Alb3 und cpSRP43.........................86 4.2.8 Untersuchung der Alb3 Bindestelle im Thylakoidmembran-Bindungstest.....89 4.2.9 Analyse der Bindedomäne des cpSRP43 für das Alb3.................................90 4.3 Analyse der Interaktion von Alb3 und Alb4 mit den Proteinen der
Thylakoidmembran........................................................................................95 4.3.1 Crosslinking der Thylakoidmembranproteine................................................95 4.3.2 Interaktion von Alb4 mit den kernkodierten ATP-Synthase
5.1 Die cpSRP-Untereinheit cpSRP43 interagiert direkt mit dem integralen Membranprotein Alb3..................................................................................103
5.2 Identifizierung der Alb3-Interaktionsbereiche für das cpSRP43 und Charakterisierung der Alb3-cpSRP43 Interaktion .......................................105
5.3 Das cpSRP43 interagiert mit Alb3 über die Ankyrin repeat Region ............110 5.4 Alb4 interagiert nicht mit den Komponenten des cpSRP-Transportwegs ...115 5.5 Alb4 ist mit der chloroplastidären ATP-Synthase assoziiert und interagiert
nicht mit Alb3 und cpSecY ..........................................................................116
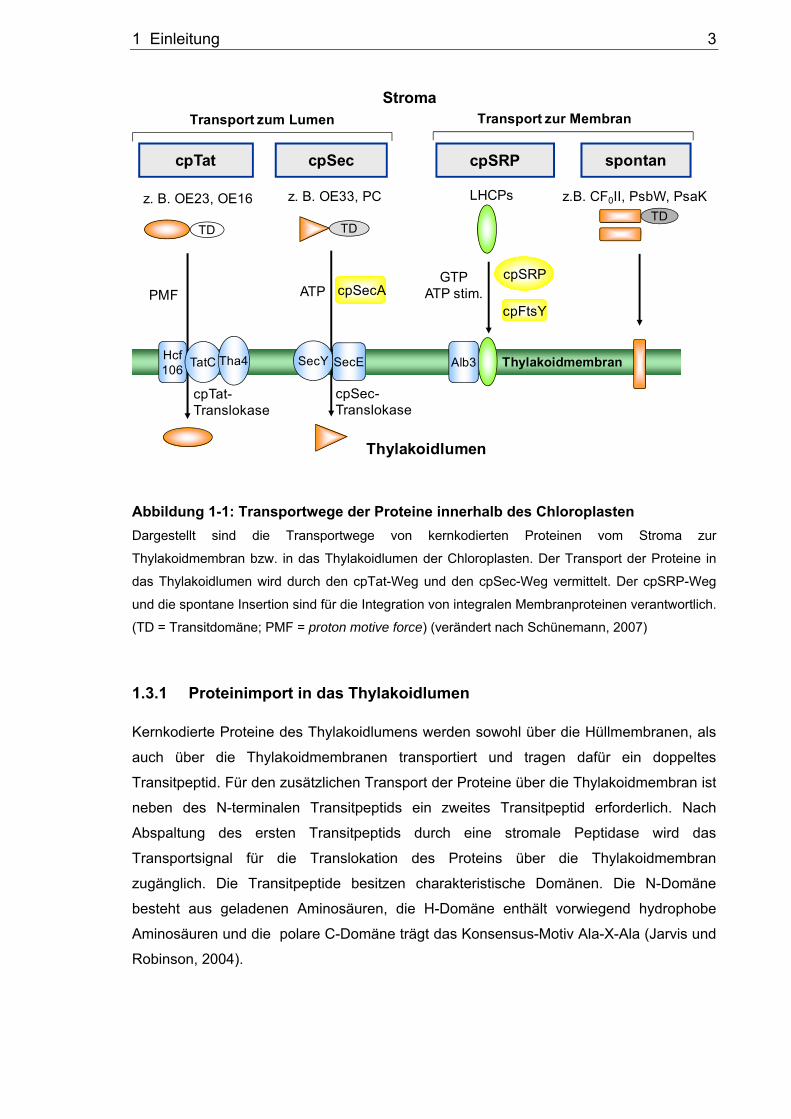
Kernkodierte Proteine des Thylakoidlumens werden sowohl über die Hüllmembranen, als
auch über die Thylakoidmembranen transportiert und tragen dafür ein doppeltes
Transitpeptid. Für den zusätzlichen Transport der Proteine über die Thylakoidmembran ist
neben des N-terminalen Transitpeptids ein zweites Transitpeptid erforderlich. Nach
Abspaltung des ersten Transitpeptids durch eine stromale Peptidase wird das
Transportsignal für die Translokation des Proteins über die Thylakoidmembran
zugänglich. Die Transitpeptide besitzen charakteristische Domänen. Die N-Domäne
besteht aus geladenen Aminosäuren, die H-Domäne enthält vorwiegend hydrophobe
Aminosäuren und die polare C-Domäne trägt das Konsensus-Motiv Ala-X-Ala (Jarvis und
Robinson, 2004).
1 Einleitung 4
1.3.1.1 Der cpSec-Weg
Der chloroplastidäre Sec-Weg zeigt Homologien zum bakteriellen Transportmechanismus
von Sec-abhängigen Proteinen (Robson und Collinson, 2006; Driessen und Nouwen,
2008). In Bakterien bindet das cytosolische Chaperon SecB das Vorstufenprotein und
transportiert dieses zur periplasmatischen Membran. Das periphere Membranprotein
SecA bildet als eine ATPase den Translokationsmotor und schleust die Proteine durch
den Translokationskanal. Die Translokationspore wird von den essentiellen
Hauptkomponenten SecY und SecE gebildet (Brundage et al., 1990). Der cotranslationale
Transport erfolgt über das cytosolische SRP.
Der bakterielle und chloroplastidäre Sec-abhängige Proteintransport erfolgt nach
scheinbar ähnlichen Mechanismen. Die Proteine werden ebenfalls im ungefalteten
Zustand transportiert. Es konnten bereits mit der ATPase cpSecA und den Proteinen der
Translokase cpSecY und cpSecE chloroplastidäre Homologe nachgewiesen werden
(Abbildung 1-1) (Yuan et al., 1994; Laidler et al., 1995; Roy und Barkan, 1998;
Schünemann et al., 1999; Sun et al., 2007). Jedoch konnten nicht zu allen bakteriellen
Komponenten homologe Proteine im chloroplastidären cpSec-Transport gefunden
werden. Aufgrund der Assoziation des YidC-Homologs Alb3 (siehe auch Abschnitt 1.5)
und des chloroplastidär kodierten D1 mit cpSecY wird derzeit eine Funktion der cpSec-
Translokase bei der cotranslationalen Insertion von Proteinen in die Thylakoidmembran
diskutiert (Zhang et al., 2001; Klostermann et al., 2002; Pasch et al., 2005).
1.3.1.2 Der cpTat-Weg
Eine weitere Möglichkeit für die Translokation von lumenalen Proteinen bietet der cpTat-
Transportweg (Abbildung 1-1). Die Transitpeptide der Proteine, die über diesen Weg
transportiert werden, tragen ein charakteristisches Twin-arginine-Motiv, welches diesem
Transportweg seinen Namen verleiht (Tat: Twin-arginine translocation) (Chaddock et al.,
1995). Eine Besonderheit des cpTat-Wegs ist, dass bereits gefaltene Proteine über die
Thylakoidmembran translokalisiert werden können (Marques et al., 2004). Für diesen
Transport sind im Gegensatz zur cpSec-Translokase keine löslichen Komponenten
bekannt. Für den Translokationsvorgang sind die integralen Membranproteine Hcf106,
Tha4 und cpTatC essentiell (Cline und Mori, 2001; Mori und Cline, 2002; Dabney-Smith et
al., 2006). Die Untereinheiten Hcf106 und cpTatC bilden einen Rezeptorkomplex,
während das Tha4 Protein durch Oligomerisierung einen Kanal formt. Bei einer Bindung
des Substratproteins kommt es zur Assemblierung der beiden Komplexe zur vollständigen
Translokase.
Der Translokationsvorgang erfordert in vitro einen Protonengradienten (Klösgen et al.,
1 Einleitung 5
1992), während in vivo diese Abhängigkeit nicht bestätigt werden konnte (Finazzi et al.,
2003; Di Cola et al., 2005). Dieses könnte an fehlenden Komponenten in den in vitro
Studien liegen. Anstelle dessen wird eine Bedeutung des Membranpotentials diskutiert
(Theg et al., 2005; Braun et al., 2007).
1.3.2 Proteinintegration in die Thylakoidmembran
1.3.2.1 Spontane Insertion
Die meisten chloroplastidären Proteine werden anhand eines spezifischen Transportwegs
über die Thylakoidmembran translokalisiert bzw. in diese inseriert. Einige Proteine jedoch
werden offenbar ohne Beteiligung von Energiekomponenten, stromalen
Transportproteinen oder einer Translokase in die Thylakoidmembran integriert (Abbildung
1-1) (Michl et al., 1994; Kim et al., 1999; Woolhead et al., 2001). Die Integration dieser
Proteine scheint sich ohne irgendwelche Hilfskomponenten auf spontanem Weg zu
vollziehen. Über die spontane Insertion werden hauptsächlich Proteine mit einer
Transmembrandomäne (CF0II, PsbX, PsbW) in die Membran eingebaut. Bei diesen
Proteinen bedingt eine hydrophobe Region des Transitpeptids die Insertion (Thompson et
al., 1998). Allerdings kann die spontane Insertion auch von Proteinen mit zwei
Transmembrandomänen vollzogen werden (PsaK, PsaG). In diesem Fall ermöglicht eine
positiv geladene stromale loop Region die Insertion der Proteine in die Thylakoidmembran
(Zygadlo et al., 2006).
1.3.2.2 Der cpSRP-Weg
Die SRPs (signal recognition particles) sind aus dem Cytosol von Eukaryoten und
Prokaryoten bekannt, wo sie der cotranslationalen Insertion von Membranproteinen in das
endoplasmatische Retikulum und in die Plasmamembran dienen (Koch et al., 2003; Pool,
2005). Das SRP ist ein Ribonukleoprotein, welches bei Eukaryoten aus einer RNA-
Komponente und sechs Polypeptiden besteht. Die etwa 54 kDa große Untereinheit
SRP54 erkennt das Signalpeptid des zu transportierenden Proteins. Nach Bindung dieses
Komplexes an den SRP-Rezeptor wird das Protein cotranslational durch einen
Translokationskanal in das endoplasmatische Retikulum geschleust.
Der SRP-Weg in E. coli ähnelt dem eukaryotischen SRP-Weg. Der SRP-
Transportmechanismus dient dem cotranslationalen Transport von Proteinen zur
Plasmamembran (de Gier und Luirink, 2001). Das SRP in E. coli stellt mit einer RNA-
1 Einleitung 6
Komponente und dem bakteriellen Homolog des SRP54, Ffh (fiftyfour homolog), ebenfalls
ein Ribonukleoprotein dar. Bei dem FtsY handelt es sich um einen SRP-Rezeptor. Dieser
Rezeptor ist als peripheres Protein der Plasmamembran und als lösliches Protein im
Cytosol zu finden. Das cytosolische FtsY bindet den Komplex aus Ribosom, SRP und
naszierendem Protein und befördert diesen zur Plasmamembran.
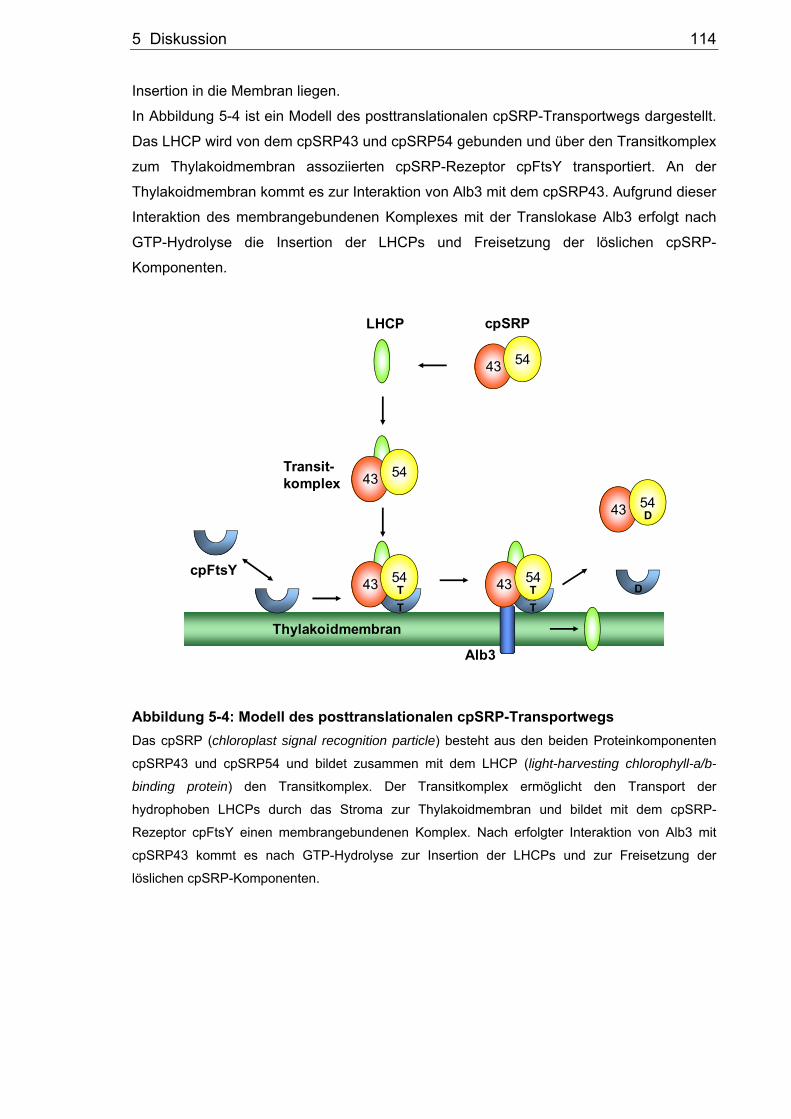
Das chloroplastidäre SRP kann im Vergleich zu den anderen SRPs sowohl
cotranslational, als auch posttranslational wirken. Der posttranslationale chloroplastidäre
SRP-Weg (cpSRP-Weg) dient der Integration der Proteine der LHCP-Familie (light-
harvesting chlorophyll a/b-binding protein) in die Thylakoidmembran (Abbildung 1-1)
(Schünemann, 2007). Die LHCPs agieren als periphere Antennen der Photosysteme I und
II und stellen etwa ein Drittel der gesamten Thylakoidmembranproteine (Yamamoto und
Bassi, 1996). Bei den LHCPs handelt es sich um kernkodierte Proteine, die als
Vorstufenproteine im Cytosol synthetisiert werden. Ein N-terminales Transitpeptid sorgt
für den posttranslationalen Transport über das TOC- und TIC-Translokon der
chloroplastidären Hüllmembranen ins Stroma. Nachdem das Transitpeptid abgespalten
wurde, werden die LHCPs durch das chloroplastidäre SRP gebunden (Abbildung 1-2).
Der so geformte Transitkomplex ermöglicht den Transport der hydrophoben LHCPs durch
das Stroma zur Thylakoidmembran und hält das LHCP somit in einem integrationsfähigen
Zustand (Payan und Cline, 1991). Chloroplastidäre SRPs beinhalten keine RNA, besitzen
jedoch neben einem SRP54-Homolog (cpSRP54) noch eine weitere 43 kDa große
Komponente (cpSRP43) (Li et al., 1995; Schünemann et al., 1998; Klimyuk et al., 1999).
Bei dem cpSRP43 handelt es sich um eine neue Komponente der SRP-Untereinheiten,
die bisher nur in Chloroplasten Höherer Pflanzen identifiziert wurde. Allerdings konnte von
unserer Arbeitsgruppe gezeigt werden, dass es ein Homolog zum cpSRP43 in
Chlamydomonas reinhardtii und Physcomitrella patens gibt (Richter, Träger und
Schünemann, unveröffentlichte Daten). Der chloroplastidäre SRP-Rezeptor cpFtsY ist
sowohl im Stroma lokalisiert, als auch mit der Thylakoidmembran assoziiert (Kogata et al.,
1999; Tu et al., 1999). Analog zum bakteriellen System scheint das cpFtsY als Rezeptor
des Transitkomplexes zu arbeiten, der für den Transport zum Translokon der Membran
verantwortlich ist. Des Weiteren spielen die Nukleotide ATP und GTP eine entscheidende
Rolle in diesem Transportweg (Hoffman und Franklin, 1994; Yuan et al., 2002). Die
Funktion des ATP ist allerdings noch nicht geklärt. Bisher wurden nur GTPasen
identifiziert. Die GTP-Hydrolyse der GTPasen cpSRP54 und cpFtsY ermöglicht einen
effizienten und gerichteten Transport (Jaru-Ampornpan et al., 2007). Es wurde gezeigt,
dass für die in vitro Integration der LHCPs in isolierte Thylakoidmembranen die
Komponenten cpSRP43, cpSRP54, cpFtsY und GTP ausreichend sind (Tu et al., 1999;
Yuan et al., 2002). Die detaillierten Proteininteraktionen der einzelnen Komponenten des
posttranslationalen cpSRP-Transportwegs sind einem folgenden Kapitel zu entnehmen
1 Einleitung 7
(Kapitel 1.4). Mit dem integralen Thylakoidmembranprotein Alb3 wurde die Translokase
des posttranslationalen cpSRP-Wegs entdeckt, die für die Insertion der LHCPs in die
Thylakoidmembran notwendig ist (Sundberg et al., 1997; Moore et al., 2000). Das Alb3
gehört zu der sogenannten Oxa1/YidC/Alb3- Proteinfamilie, dessen Mitglieder in der
Insertion und Assemblierung von Membranproteinen in Mitochondrien, Bakterien und
Chloroplasten beteiligt sind (Yi und Dalbey, 2005; Xie und Dalbey, 2008). Eine
ausführliche Darstellung dieser wichtigen Membranproteine erfolgt in einem gesonderten
Kapitel zur Oxa1/YidC/Alb3- Proteinfamilie (Kapitel 1.5).
4354
cpSRP
Transit-komplex
+ GTP
LHCP
Alb3
TM
4354psbA
Ribosom
D1
cpSec-Translokase
54
54
cpFtsY
cotranslational posttranslational
Abbildung 1-2: Modell des co- und posttranslationalen cpSRP-Transportwegs Das Modell stellt die Beteiligung des cpSRP54 am cotranslationalen SRP-Weg zur Integration von
plastidär kodiertem D1 und am posttranslationalen SRP-Weg zur Integration von kernkodiertem
LHCP dar. Das cpSec-assoziierte Alb3 scheint an der SRP-abhängigen cotranslationalen Insertion
von plastidär kodierten Thylakoidmembranproteinen beteiligt zu sein, während die
posttranslationale Integration von LHCP unabhängig von der cpSec-Translokase verläuft.
(verändert nach Schünemann, 2007)
Das cpSRP54 ist neben dem posttranslationalen Transport im Komplex mit cpSRP43
auch am cotranslationalen Transport chloroplastidär kodiertem D1-Protein involviert
(Abbildung 1-2), wobei cpSRP54 an das naszierende D1 bindet (Nilsson et al., 1999;
1 Einleitung 8
Nilsson und van Wijk, 2002). Eine Interaktion des cpSRP43 mit dem naszierenden D1
konnte bislang nicht gezeigt werden. Dieses weist auf eine spezialisierte Rolle des
cpSRP43 im posttranslationalen Transport und eine Doppelfunktion des cpSRP54 sowohl
im posttranslationalen als auch im cotranslationalen Transport hin. Des Weiteren konnte
eine Assoziation von cpSecY mit Ribosomen und dem D1-Protein gezeigt werden (Zhang
et al., 2001). Die cpSec-Translokase ist somit wahrscheinlich an der cotranslationalen
Insertion des chloroplastidär kodierten D1 beteiligt. Das integrale Membranprotein Alb3
scheint ebenfalls eine Doppelfunktion innerhalb des cpSRP-Wegs zu haben. Neben der
Funktion in der posttranslationalen Insertion der LHCPs in die Thylakoidmembran, scheint
die Assoziation mit der cpSec-Translokase auf eine Rolle von Alb3 innerhalb der
cotranslationalen Insertion von Proteinen zu deuten (Klostermann et al., 2002; Pasch et
al., 2005).
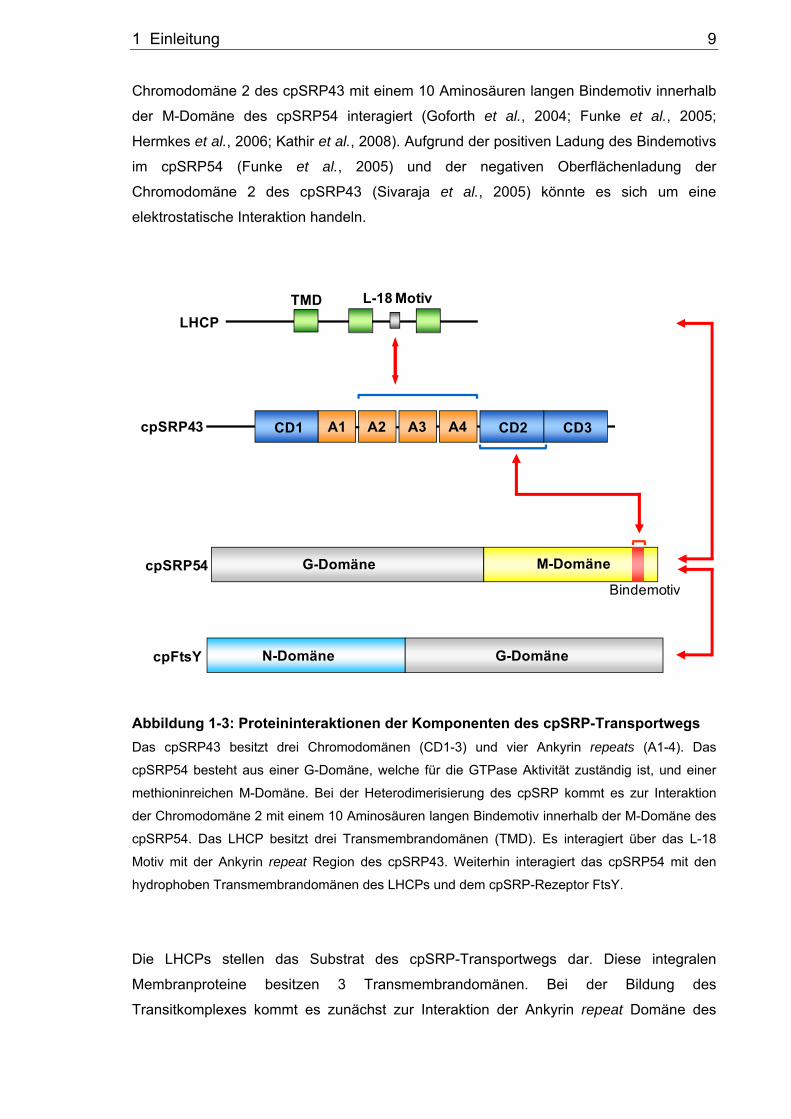
1.4 Proteininteraktionen der Komponenten des posttranslationalen cpSRP-Transportwegs
Im folgenden Abschnitt sollen die Proteininteraktionen innerhalb des cpSRP-Wegs näher
beschrieben werden (Abbildung 1-3). Das cpSRP43 ist aus zwei charakteristischen
Protein-Protein-Interaktionsdomänen aufgebaut, den Ankyrin repeats und den
Chromodomänen. Es besitzt im N-Terminus eine Chromodomäne und vier Ankyrin
repeats sowie im C-terminalen Bereich zwei benachbarte Chromodomänen (Klimyuk et
al., 1999; Jonas-Straube et al., 2001; Goforth et al., 2004). Ankyrin repeats ermöglichen
als charakteristische Bindedomänen verschiedene Proteininteraktionen und sind dadurch
an der Ausbildung von Proteinkomplexen beteiligt (Sedgwick und Smerdon, 1999). Die
Chromodomänen wurden anfänglich in Proteinen des Zellkerns identifiziert, wo sie einen
Einfluss auf die Regulation der Chromatinstruktur zeigen (Brehm et al., 2004).
Chromodomänen sind charakteristisch aus drei antiparallelen β-Faltblättern und einer C-
terminalen α-Helix aufgebaut (Sivaraja et al., 2005). Die Kristallstruktur des cpSRP43
(Chromodomäne1 bis Ankyrin repeat 4) zeigt, dass die Ankyrin repeats 2 und 3 ein
typisches helix-turn-helix Motiv besitzen, während die Ankyrin repeats 1 und 4 verlängerte
Helices aufweisen (Stengel et al., 2008). Die Chromodomäne 1 und Ankyrin repeat 1
gehen durch Fusion ihrer Helices direkt ineinander über. Auch Ankyrin repeat 4 zeigt
durch die verlängerte Helix einen direkten Übergang zur Chromodomäne 2. Das cpSRP54
besteht aus einer N-terminalen G-Domäne, welche für die GTPase Funktion
verantwortlich ist, und einer C-terminalen methioninreichen M-Domäne (Franklin und
Hoffman, 1993). Bei der Heterodimerisierung des cpSRP-Komplexes kommt es zur
Interaktion der Chromodomäne 2 des cpSRP43 mit der M-Domäne des cpSRP54
(Goforth et al., 2004; Hermkes et al., 2006). Es konnte gezeigt werden, dass die
1 Einleitung 9
Chromodomäne 2 des cpSRP43 mit einem 10 Aminosäuren langen Bindemotiv innerhalb
der M-Domäne des cpSRP54 interagiert (Goforth et al., 2004; Funke et al., 2005;
Hermkes et al., 2006; Kathir et al., 2008). Aufgrund der positiven Ladung des Bindemotivs
im cpSRP54 (Funke et al., 2005) und der negativen Oberflächenladung der
Chromodomäne 2 des cpSRP43 (Sivaraja et al., 2005) könnte es sich um eine
elektrostatische Interaktion handeln.
LHCPTMD L-18 Motiv
cpSRP43 A1 A2 A3 A4 CD3
cpSRP54 G-Domäne M-Domäne
Bindemotiv
cpFtsY G-DomäneN-Domäne
CD2CD2CD1
Abbildung 1-3: Proteininteraktionen der Komponenten des cpSRP-Transportwegs Das cpSRP43 besitzt drei Chromodomänen (CD1-3) und vier Ankyrin repeats (A1-4). Das
cpSRP54 besteht aus einer G-Domäne, welche für die GTPase Aktivität zuständig ist, und einer
methioninreichen M-Domäne. Bei der Heterodimerisierung des cpSRP kommt es zur Interaktion
der Chromodomäne 2 mit einem 10 Aminosäuren langen Bindemotiv innerhalb der M-Domäne des
cpSRP54. Das LHCP besitzt drei Transmembrandomänen (TMD). Es interagiert über das L-18
Motiv mit der Ankyrin repeat Region des cpSRP43. Weiterhin interagiert das cpSRP54 mit den
hydrophoben Transmembrandomänen des LHCPs und dem cpSRP-Rezeptor FtsY.
Die LHCPs stellen das Substrat des cpSRP-Transportwegs dar. Diese integralen
Membranproteine besitzen 3 Transmembrandomänen. Bei der Bildung des
Transitkomplexes kommt es zunächst zur Interaktion der Ankyrin repeat Domäne des
1 Einleitung 10
cpSRP43 mit dem sogenannten L18-Motiv des LHCPs, welches sich zwischen der 2. und
3. Transmembrandomäne befindet (DeLille et al., 2000; Tu et al., 2000; Jonas-Straube et
al., 2001; Stengel et al., 2008). Die Bindung zwischen cpSRP54 und LHCP erfolgt über
die hydrophoben Transmembrandomänen des LHCPs (DeLille et al., 2000; Tu et al.,
2000; Groves et al., 2001). Das cpFtsY ist aus einer N-terminalen N-Domäne und einer C-
terminalen G-Domäne, welche für die GTPase Aktivität verantwortlich ist, aufgebaut
(Stengel et al., 2007; Chandrasekar et al., 2008). Die Proteine cpSRP54 und cpFtsY sind
in der Lage in GTP-gebundener Konformation miteinander zu interagieren. Die
Dissoziation beider Proteine erfolgt über GTP-Hydrolyse (Jaru-Ampornpan et al., 2007).
Das Protein Alb3 stellt die Translokase des cpSRP-Wegs dar. Dieses integrale
Membranprotein besitzt 5 Transmembrandomänen. Es konnte gezeigt werden, dass
cpSRP54 und cpFtsY zusammen mit Alb3 einen Komplex an der Thylakoidmembran
bilden können (Moore et al., 2003). Die Proteininteraktionen der löslichen cpSRP-
Komponenten mit Alb3 sind jedoch noch nicht vollständig geklärt und daher ein zentraler
Teil dieser Arbeit.
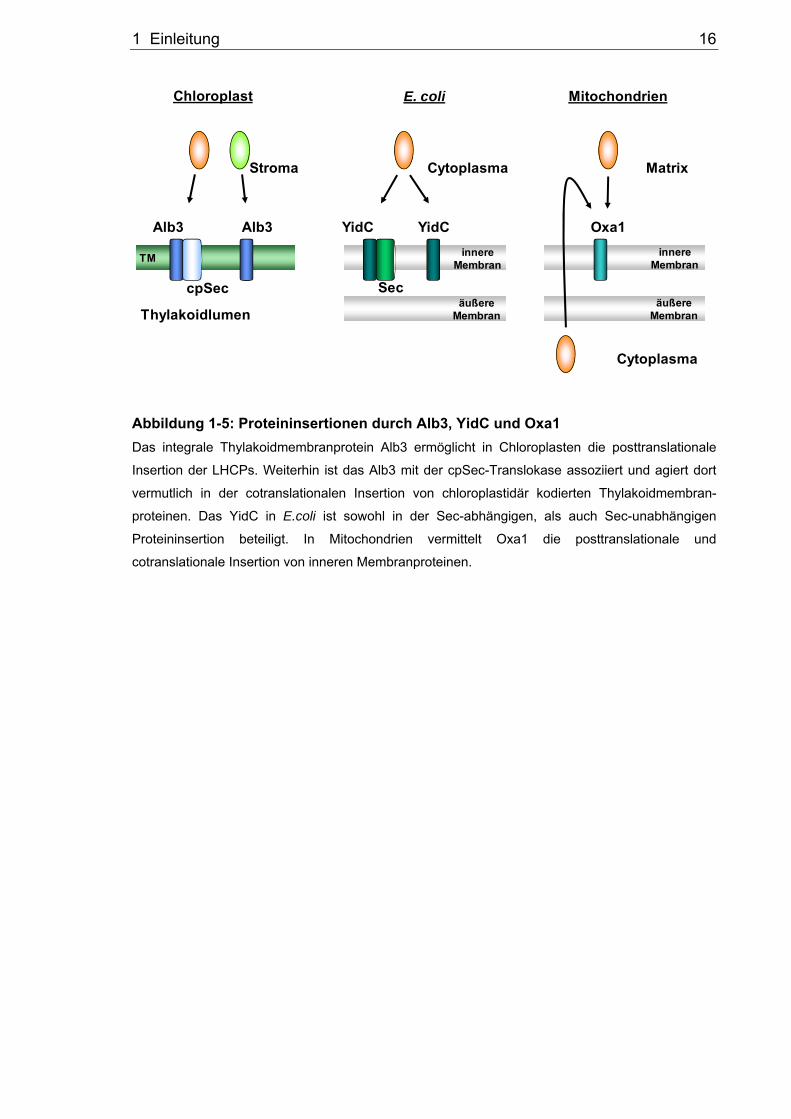
1.5 Die Oxa1/YidC/Alb3-Proteinfamilie
Die Biogenese von membrangebundenen Proteinkomplexen wie z.B. der Photosysteme,
Atmungskette oder ATP-Synthase ist von entscheidender Bedeutung für die Funktionalität
von Zellen. Für diese Funktionalität der Proteinkomplexe ist nicht nur der Transport der
Proteine an ihren Wirkort, sondern auch ein korrekter Einbau in die Membranen essentiell.
Die Mitglieder der sogenannten Oxa1/YidC/Alb3-Proteinfamilie sind evolutionär
konserviert und ermöglichen die Insertion und Assemblierung von Membranproteinen in
Mitochondrien, Bakterien und Chloroplasten (Kuhn et al., 2003; Kiefer und Kuhn, 2007;
Xie und Dalbey, 2008; Zhang et al., 2009). Diese Proteinfamilie weist im Bereich ihrer
Transmembrandomänen eine hydrophobe Kernregion auf, während die hydrophilen
Bereiche in ihrer Länge und Funktion variieren (Yen et al., 2001). Die konservierten
Bereiche der Proteine ermöglichen die Komplementation der Proteine untereinander in
den verschiedenen Systemen, während die hydrophilen Anteile auf eine spezielle
Funktion der Proteine deuten (Preuss et al., 2005; van Bloois et al., 2005). Das bakterielle
Homolog YidC besitzt 6 Transmembrandomänen, während die Homologen in den
Organellen, Oxa1 und Alb3, jeweils 5 Transmembrandomänen aufweisen (Abbildung 1-4).
Die N- und C-terminalen Bereiche des bakteriellen YidC ragen, aufgrund der
Membrantopologie, beide ins Cytoplasma. Das YidC besitzt zwischen der ersten und
zweiten Transmembrandomäne eine lange periplasmatische Domäne. Erste Arbeiten
1 Einleitung 11
geben näheren Aufschluss auf die Struktur des YidC (Ravaud et al., 2008). Bei dem
mitochondrialen Oxa1 und dem chloroplastidären Alb3 ergibt die Membrantopologie eine
Lokalisation der N- und C-terminalen Enden auf gegenüberliegenden Seiten. Der
N-Terminus von Oxa1 zeigt in den Intermembranraum und der C-Terminus in die Matrix
der Mitochondrien. Das chloroplastidäre Alb3 ragt mit dem N-Terminus in das
Thylakoidlumen und mit dem C-terminalen Ende in das Stroma (Kuhn et al., 2003). In den
folgenden Abschnitten sollen zunächst die charakteristischen Funktionen der
namensgebenden Mitglieder der Oxa1/YidC/Alb3-Proteinfamilie näher beschrieben
werden. Zusätzlich erfolgt eine Beschreibung weiterer Homologe dieser Proteinfamilie.
N
C
N
N CC
YidC Oxa1Alb3
Intermembranraum
MatrixCytoplasma
Thylakoidlumen
Stroma
Periplasma
Abbildung 1-4: Membrantopologie von Alb3, YidC und Oxa1 Das chloroplastidäre Alb3 und das mitochondriale Oxa1 besitzen fünf Transmembrandomänen,
während das bakterielle Homolog YidC aus sechs Transmembrandomänen besteht (Kuhn et al.,
2003). Die N- und C-terminalen Bereichen des Alb3 und Oxa1 befinden sich auf
gegenüberliegenden Seiten der Membran. Der N-Terminus von Alb3 zeigt ins Thylakoidlumen und
der C-Terminus ins Stroma der Chloroplasten. Der N-Terminus des mitochondrialen Oxa1 weist in
den Intermembranraum und der C-Terminus in die Matrix der Mitochondrien. Das bakterielle YidC
zeigt eine Membrantopologie, bei der beide Termini ins Cytoplasma ragen. Das YidC besitzt
zwischen der ersten und zweiten Transmembrandomäne eine lange periplasmatische Domäne.
Oxa1 und Alb3 haben jeweils einen verlängerten hydrophilen C-Terminus.
1.5.1 Oxa1
Oxa1 ist ein 36 kDa großes Protein der inneren Mitochondrienmembran von
Saccharomyces cerevisiae, das die Insertion von kern- und mitochondrialkodierten,
inneren Membranproteinen vermittelt (Hell et al., 1998; Hell et al., 2001) (Abbildung 1-5).
Es wurde durch die Analyse von Proteinen, die an der Assemblierung des Cytochrom-
1 Einleitung 12
Oxidase-Komplexes beteiligt sind, identifiziert (Oxa1 = oxidase assembly 1) (Bauer et al.,
1994; Bonnefoy et al., 1994). Oxa1 ist sowohl an der posttranslationalen, als auch an der
cotranslationalen Proteininsertion beteiligt (Bonnefoy et al., 2009). Im mitochondrialen
Transport sind bislang keine Homologen zu den Untereinheiten der bakteriellen Sec-
Translokase bekannt. Daher scheint das Oxa1 ausschließlich in einem Sec-unabhängigen
Integrationsmechanismus zu agieren (Glick und Von Heijne, 1996). Für die Oxa1-
abhängige Insertion von kernkodierten, mitochondrialen Proteinen werden die Proteine
zunächst vom Cytosol in die mitochondriale Matrix transportiert und anschließend in die
innere Membran integriert. Durch Interaktion der C-terminalen, α-helicalen Domäne des
Oxa1 mit den translatierenden Ribosomen konnte eine unterstützende Wirkung auf die
cotranslationale Proteininsertion nachgewiesen werden (Jia et al., 2003; Szyrach et al.,
2003). Diese C-terminale Domäne scheint jedoch keinen Einfluss auf die
posttranslationale Proteininsertion zu haben. Es konnte gezeigt werden, dass das
bakterielle Homolog YidC mit Hilfe der C-terminalen Ribosomen-Bindestelle des Oxa1 die
Funktion des Oxa1 komplementieren kann (Preuss et al., 2005; van Bloois et al., 2005).
Weiterhin wurde gezeigt, dass Oxa1 wie auch das bakterielle Homolog YidC mit
translatierenden Ribosomen eine Pore bildet (Kohler et al., 2009). Oxa1 Hefemutanten
haben gezeigt, dass von der Deletion des Oxa1 nicht nur die Assemblierung der
Cytochrom-Oxidase, sondern auch die ATP-Synthase betroffen ist (Altamura et al., 1996;
Jia et al., 2007).
In Mitochondrien von Saccharomyces cerevisiae wurde neben dem Oxa1 bereits ein
Homolog identifiziert, welches als Oxa2/Cox18 bezeichnet wurde. Dieses Protein spielt
eine spezifische Rolle in der Biogenese des Cytochrom-Oxidase-Komplexes (Preuss et
al., 2005; Bonnefoy et al., 2009). Komplementationsversuche haben gezeigt, dass YidC
die beiden mitochondrialen Homologe Oxa1 und Oxa2/Cox18 funktionell ersetzen kann
(Preuss et al., 2005). Das Oxa2/Cox18 kann ebenfalls die Sec-unabhängige Funktion von
YidC in E. coli komplementieren (van Bloois et al., 2007). In der Hefe sind Oxa1 und
Oxa2/Cox18 nicht in der Lage sich gegenseitig zu komplementieren, was auf eine
unterschiedliche Funktion der beiden Proteine schließen lässt. Das Oxa2/Cox18 scheint
im Gegensatz zum Oxa1 ausschließlich eine posttranslationale Aufgabe zu haben. Die
posttranslationale Funktion des Oxa2/Cox18 wird durch die unterstützende Wirkung auf
die Translokation der C-terminalen Domäne des Cox2 in den Intermembranraum und eine
mögliche Funktion in der Assemblierung mit den anderen Untereinheiten des Cytochrom-
Oxidase-Komplexes gezeigt (Saracco und Fox, 2002). Das Oxa2 in Neurospora crassa ist
in der Lage die Oxa2-Mutante der Hefe zu komplementieren, was für eine gemeinsame
Funktion der beiden Proteine spricht. Die Deletion dieses Oxa2-Gens verursacht einen
spezifischen Defekt in der Biogenese des Cytochrom-Oxidase-Komplexes (Funes et al.,
2004).
1 Einleitung 13
1.5.2 Yid C
YidC ist ein 60 kDa großes, integrales Membranprotein der Plasmamembran in E. coli und
wurde als neue Komponente der Sec-Translokase nachgewiesen (Saaf et al., 1998; Scotti
et al., 2000). In E. coli werden die meisten Membranproteine über den cotranslationalen
SRP-Weg zur Plasmamembran transportiert und über die Sec-Translokase in die innere
Membran inseriert. Das integrale Membranprotein der Plasmamembran YidC stellt eine
entscheidende Komponente der Sec-Translokase dar und ermöglicht den lateralen
Einbau von Transmembrandomänen integrierter Proteine aus der Sec-Translokase in die
Membran (Beck et al., 2001; Urbanus et al., 2001; van der Laan et al., 2001; Houben et
al., 2004). YidC interagiert dabei mit den Transmembrandomänen der inserierten Proteine
direkt nach deren Freisetzung aus dem Sec-Translokon. Dieses verhindert die
Aggregation der integrierten Proteine. Das YidC scheint somit eine Funktion als
Membranchaperon auszuüben. Des Weiteren wurde gezeigt, dass YidC in E. coli für die
Insertion von Membranproteinen, die unabhängig von der Sec-Translokase integriert
werden, essentiell ist (Samuelson et al., 2000; Chen et al., 2002; Serek et al., 2004; van
der Laan et al., 2004; van Bloois et al., 2005; Facey et al., 2007). Dabei handelt es sich
z. B. um Phagenproteine oder die F0c-Untereinheit der ATP-Synthase. Von den
Phagenproteinen wurde zunächst angenommen, dass sie spontan in die Membran
inserieren. Das YidC ist in der Lage, wie das mitochondriale Homolog Oxa1, mit
translatierenden Ribosomen eine dimere Insertionspore auszubilden (Kohler et al., 2009).
Die bisherigen Daten zeigen, dass YidC sowohl in Kombination mit der Sec-Translokase
als auch unabhängig von dieser bei der Integration von Membranproteinen agieren kann
(Abbildung 1-5).
In grampositiven Bakterien findet man hauptsächlich zwei YidC Homologe (Funes et al.,
2009). Das YidC1 und YidC2 aus Streptococcus mutans sind in der Lage das YidC aus
E.coli in der Sec-abhängigen und Sec-unabhängigen Proteininsertion zu komplemetieren.
YidC2 besitzt, wie das mitochondriale Homolog Oxa1, einen positiv geladenen C-
Terminus und kann mit Ribosomen assoziieren. Es ist in der Lage die Funktion von Oxa1
in der cotranslationalen Proteininsertion von mitochondrialen, inneren Membranproteinen
zu übernehmen. Weiterhin wurde gezeigt, dass YidC2 unabhängig von dem SRP-Weg
cotranslational wirken kann.
1 Einleitung 14
1.5.3 Alb3
Das Alb3 Protein wurde aus der Analyse von Pigment-defizienten Mutanten von
Arabidopsis thaliana identifiziert. Die Mutanten zeigen einen drastischen Albino-Phänotyp
(Albino 3 = Alb3) und sind nicht in der Lage ohne externe Kohlenstoffquelle über das
Keimungsstadium hinaus zu überleben (Sundberg et al., 1997). Der Chlorophyllgehalt
beträgt nur etwa 5 % des Wildtyplevels. Die Chloroplasten zeigen nur wenige
Thylakoidmembranen und keinerlei Granastapelung. Alb3 ist ein 45 kDa großes,
integrales Thylakoidmembranprotein, welches an der posttranslationalen Integration der
LHCPs in die Thylakoidmembran beteiligt ist (Moore et al., 2000; Moore et al., 2003)
(Abbildung 1-5). Es konnte gezeigt werden, dass eine Vorinkubation von
Thylakoidmembranen mit einem Alb3-Antikörper die Integration des Lhcb1 beeinträchtigt,
während dieses keine Auswirkungen auf den Transport von cpSec- und cpTat-
abhängigen Proteinen hatte (Moore et al., 2000). Gleichermaßen wurde die Integration
von zwei weiteren Mitgliedern der LHCP-Familie (Lhcb4.1 und Lhcb5) in die
Thylakoidmembran verhindert (Woolhead et al., 2001). Das Alb3 scheint bei der
Integration von LHCP unabhängig von der cpSec-Translokase zu agieren, da die
Vorinkubation der Thylakoidmembran mit einem Antikörper gegen den C-Terminus von
cpSecY keinen Einfluss auf die Integration von LHCP hat (Mori et al., 1999). Die
posttranslationale Integration der LHCPs ist daher ein cpSRP/Alb3-abhängiger, jedoch
cpSec-unabhängiger Mechanismus (Moore et al., 2003). Andere Arbeiten zeigen
allerdings eine Assoziation des Alb3 mit der cpSec-Translokase (Klostermann et al., 2002;
Pasch et al., 2005). Das cpSec-assoziierte Alb3 scheint an der cpSRP-abhängigen
cotranslationalen Insertion von chloroplastidär kodierten Thylakoidmembranproteinen
beteiligt zu sein (Abbildung 1-5). Zudem konnte gezeigt werden, dass Alb3 in der Lage ist
das YidC in einem YidC-defizienten Bakterienstamm zu komplementieren. Das Alb3
ermöglicht in E.coli die Membraninsertion von Sec-abhängigen und Sec-unabhängigen
Membranproteinen (Jiang et al., 2002).
Neben dem Alb3 konnte mit dem Alb4 ein zweites chloroplastidäres Homolog der
Oxa1/YidC/Alb3-Proteinfamilie in Arabidopsis thaliana identifiziert werden (Gerdes et al.,
2006). Zahlreiche Analysen wie z. B. Importstudien, immunologische Detektion in
chloroplastidären Subfraktionen und GFP-Lokalisationsstudien ergaben, dass Alb4 in der
Thylakoidmembran der Chloroplasten lokalisiert ist. Arabidopsis thaliana Mutanten mit
einem um 90 % reduzierten Alb4 Level zeigten keinen offensichtlichen Phänotyp. Der im
Vergleich zu den alb4-Mutanten drastische Phänotyp der alb3-Mutante zeigt, dass Alb4
nicht in der Lage ist die Funktion von Alb3 zu kompensieren. Die chloroplastidäre
Ultrastruktur der alb4-Mutanten weist vergrößerte Plastiden mit einer im Vergleich zum
Wildtyp aufgelockerten Anordnung der Granathylakoide auf. Das Alb4 scheint somit einen
1 Einleitung 15
Einfluss auf die Biogenese der Chloroplasten zu haben. Die genaue Funktion des Alb4 ist
jedoch noch nicht bekannt.
In Chlamydomonas reinhardtii wurden ebenfalls zwei Alb3-Homologe (Alb3.1 und Alb3.2)
identifiziert (Bellafiore et al., 2002; Göhre et al., 2006). Die Inaktivierung des Alb3.1 in der
ac29-Mutante von Chlamydomonas reinhardtii hat zwei bedeutende Effekte. Die Menge
des LHCI und LHCII im Photosystem I und II ist um mehr als das 10-fache reduziert und
der Chlorophyllgehalt entspricht nur 30% des Wildtyplevels. Zudem ist das Photosystem II
um das 2-fache bzw. 10-fache in den Zellen, die in Licht bzw. bei Dunkelheit angezogen
wurden, vermindert (Bellafiore et al., 2002). Es konnte weiterhin gezeigt werden, dass
Alb3.1 an der Assemblierung des D1 in das Reaktionszentrum des Photosystems II von
Chlamydomonas reinhardtii involviert ist (Ossenbühl et al., 2004). Interessanterweise
scheint dagegen Alb3.1 für die Integration des D1 in die Thylakoidmembran keine
signifikante Rolle zu spielen (Ossenbühl et al., 2004). Das Alb3.1 hat daher eine Funktion
in der Integration der LHCPs und in der Assemblierung des Photosystem II. Weiterhin ist
bemerkenswert, dass die Inaktivierung des Alb3.1 in Chlamydomonas reinhardtii einen
deutlich milderen Phänotyp zeigt, als der Verlust des Alb3 in Arabidopsis thaliana
(Sundberg et al., 1997; Bellafiore et al., 2002). Mit dem Alb3.2 konnte ein weiteres Alb3-
Homolog in Chlamydomonas reinhardtii identifiziert werden, das den Verlust des Alb3.1 in
der Mutante ac29 möglicherweise teilweise kompensieren kann (Bellafiore et al., 2002).
Eine Reduktion des Alb3.2 in der Chlamydomonas RNAi-Mutante führt zu einer starken
Reduktion der Phtosysteme I und II und langfristig zum Zelltod. Während die Menge an
Lichtsammelkomplexen nicht so stark reduziert ist, wie in der alb3.1-Mutante, zeigt die
alb3.2-Mutante mit einer Vergrößerung der Vakuole einen weiteren Phänotyp, dessen
Ursache allerdings nicht geklärt ist (Göhre et al., 2006).
Im Cyanobakterium Synechocystis sp. PCC6803 konnte ebenfalls ein Alb3-Homolog
(Slr1471) identifiziert werden. Mutanten zeigten, dass dieses Homolog für die Morphologie
der Thylakoide und die Lebensfähigkeit der Zellen wichtig ist (Spence et al., 2004). Es
konnte gezeigt werden, dass das Slr1471 mit dem D1 Protein interagieren kann und
daher für die Integration des D1 wichtig ist (Ossenbühl et al., 2006). Das Slr1471 ist
hauptsächlich in der Cytoplasmamembran lokalisiert und in der Lage das YidC in einem
YidC-defizienten E.coli Bakterienstamm zu ersetzen (Gathmann et al., 2008). Mit der
Lokalisation des Slr1471 in der Cytoplasmamembran ist eine Funktion in der Biogenese
des Photosystem II in der Cytoplasmamembran verbunden.
1 Einleitung 16
Oxa1
Mitochondrien
Matrix
Sec
YidC
E. coli
Cytoplasma
YidC
cpSec
Alb3Alb3
Chloroplast
Thylakoidlumen
Stroma
TM innere Membran
äußere Membran
innere Membran
äußere Membran
Cytoplasma
Abbildung 1-5: Proteininsertionen durch Alb3, YidC und Oxa1 Das integrale Thylakoidmembranprotein Alb3 ermöglicht in Chloroplasten die posttranslationale
Insertion der LHCPs. Weiterhin ist das Alb3 mit der cpSec-Translokase assoziiert und agiert dort
vermutlich in der cotranslationalen Insertion von chloroplastidär kodierten Thylakoidmembran-
proteinen. Das YidC in E.coli ist sowohl in der Sec-abhängigen, als auch Sec-unabhängigen
Proteininsertion beteiligt. In Mitochondrien vermittelt Oxa1 die posttranslationale und
cotranslationale Insertion von inneren Membranproteinen.
2 Zielsetzung 17
2. Zielsetzung
In der vorliegenden Arbeit sollte die Rolle von Alb3 und Alb4 bei der Biogenese der
Thylakoidmembran von Arabidopsis thaliana genauer geklärt werden. Alb3 und Alb4 sind
chloroplastidäre Homologe der Oxa1/YidC/Alb3-Proteinfamilie, deren Mitglieder eine
entscheidende Bedeutung bei der Insertion und Assemblierung von Membranproteinen
haben.
In vorangegangenen Arbeiten konnte bereits gezeigt werden, dass Alb3 für die Insertion
von Mitgliedern der LHCPs, die cpSRP-abhängig zur Thylakoidmembran transportiert
werden, essentiell ist. In einem ersten Teilprojekt dieser Arbeit sollte der docking-Prozess
des Komplexes aus LHCP und cpSRP in Kombination mit dem SRP-Rezeptor cpFtsY an
die Thylakoidmembran näher analysiert werden. Als zentrale Frage sollte dabei
untersucht werden, ob dieser Prozess auf einer direkten Interaktion der Proteine des
cpSRP-Wegs mit der Translokase Alb3 beruht.
Da zu Beginn dieser Arbeit die molekulare Funktion des integralen
Thylakoidmembranproteins Alb4 bei der Biogenese der Chloroplasten unbekannt war,
sollte in einem weiteren Teilprojekt die Interaktion von Alb4 mit anderen
Thylakoidmembranproteinen analysiert werden.
Um die homologen Proteine in Arabidopsis thaliana in biochemischen Untersuchungen
getrennt voneinander detektieren zu können, sollten zunächst spezifische Antikörper
gegen das Alb3 und das Alb4 hergestellt werden. Für die Analyse von Protein-Protein-
Interaktionen sollten sowohl in vivo Studien unter Verwendung des Hefe Split-Ubiquitin
Systems und der bimolekularen Fluoreszenzkomplementation in Arabidopsis Protoplasten
als auch in vitro Interaktionsstudien (z. B. pull down Analysen, chemisches Crosslinking,
Coimmunpräzipitation) durchgeführt werden.
3 Material und Methoden 18
3. Material und Methoden
3.1 Chemikalien
Tabelle 3-1: Chemikalienliste
Chemikalie Hersteller [35S]-Methionin (37 TBq/mmol, 370 MBq/ml) Hartmann Adenin Hemisulfat-Salz SIGMA Affi Gel 15 Biorad Coomassie® Brilliant Blau G-250 SERVA CSM –His –Leu –Trp (Complete Supplement Mixture minus the above) BIO 101 CSM –Leu –Trp (Complete Supplement Mixture minus the above) BIO 101 Cellulase SERVA DOB (ohne AS) BIO 101 Gene RulerTM 1Kb DNA Ladder Fermentas Glutathion Sepharose 4B Amersham Macerozyme SERVA Molekulargewichtsmarker SIGMA Ni-NTA-Agarose Qiagen Page RulerTM Prestained Protein Ladder Fermentas PEG 3350 SIGMA Pepton MERCK
Ponceau S SERVA Protein A Sepharose CL 4B Amersham Proteinaseinhibitor, complete Mini Protease Inhibitor Cocktail Tablets Roche Sulfo-MBS PIERCE Super Signal West Femto Luminol/Enhancer Solution PIERCE Super Signal West Femto Stable Peroxide Buffer PIERCE Super Signal West Pico Luminol/Enhancer Solution PIERCE Super Signal West Pico Stable Peroxide Buffer PIERCE Weizenkeimlysat (Wheat Germ) Promega
Nicht aufgeführte Chemikalien wurden in Laborqualität von der Firma AppliChem
bezogen.
3 Material und Methoden 19
3.2 Verbrauchsmaterial
Tabelle 3-2: Verbrauchsmaterial
Material Hersteller Blotting Paper Schleicher&Schuell Dialyse-Kassetten PIERCE Filterpapier Schleicher&Schuell Hyperfilm ECL High performance chemiluminescence film Amersham Impfösen Greiner Kodak Biomax MR Film Scientific Imaging Film Kodak Kodak Biomax MS Scientific Imaging Film Kodak Kryoröhrchen Brand Membranfilter Millipore Miracloth Calbiochem PCR-Thermo-Strips 0,2 ml PeqLab PCR-Tubes 0,2 ml Brand Petrischalen Greiner Pipettenspitzen Greiner Polyprep Chromatographie Säulen Biorad Porablot NCL Nitrocellulosemembran Macherey-Nagel PS Küvetten halbmikro Brand Reaktionsgefäße 15 ml Greiner Reaktionsgefäße 50 ml Greiner Safelock Tubes 1,5 ml Eppendorf Safelock Tubes 2 ml Eppendorf UV-Küvetten mikro Brand Wizard Mini Columns Promega
Folgende Plasmide wurden für die in vivo Interaktionsstudien in Arabidopsis thaliana
Protoplasten eingesetzt.
pSPYNE-atpC1: for BamHI CACTAGTGGATCCATGGCTTGCTCTAATCTA rev SalI TACCCTCGAGGTCGACAACCTGTGCATTAGCTCC pSPYNE-atpC2: for BamHI CACTAGTGGATCCATGACAGGTTCGATCTCG rev SalI TACCCTCGAGGTCGACAGACTCTCGAAGAGCTTC pSPYNE-atpD: for BamHI CACTAGTGGATCCATGGCGTCTCTTCAACAA rev SalI TACCCTCGAGGTCGACAGTAGCTAATTGAATCTC pSPYNE-atpG: for BamHI CACTAGTGGATCCATGGCTGCTAATTCGATA rev SalI TACCCTCGAGGTCGACAGAAGGAAGAACCTTCTT pSPYCE-Alb4(1-142)Alb3(159-462): for BamHI CGCCACTAGTGGATCCATGTCTTCAACAATATCC for overlap GCTACGTTCCCATTGACTAAGCAACAGGTT rev overlap AACCTGTTGCTTAGTCAATGGGAACGTAGC rev SalI TACCCTCGAGGTCGACTACAGTGCGTTTCCGCTT pSPYCE-Alb4(1-210)Alb3(227-462): for BamHI CGCCACTAGTGGATCCATGTCTTCAACAATATCC for overlap CTATATAGAGCCCTCTCTAACGTGGCCAAC rev overlap GTTGGCCACGTTAGAGAGGGCTCTATATAG rev SalI TACCCTCGAGGTCGACTACAGTGCGTTTCCGCTT pSPYCE-Alb4(1-239)Alb3(256-462): for BamHI CGCCACTAGTGGATCCATGTCTTCAACAATATCC for overlap GTTGCTGCAAGACAGAGTGGATCCGGCATT rev overlap AATGCCGGATCCACTCTGTCTTGCAGCAAC rev SalI TACCCTCGAGGTCGACTACAGTGCGTTTCCGCTT
3 Material und Methoden 26
pSPYCE-Alb4(1-255)Alb3(272-462): for BamHI CGCCACTAGTGGATCCATGTCTTCAACAATATCC for overlap ATTGAGGGACACCCACCATTGGGATGGTAT rev overlap ATACCATCCCAATGGTGGGTGTCCCTCAAT rev SalI TACCCTCGAGGTCGACTACAGTGCGTTTCCGCTT pSPYCE-Alb4(1-288)Alb3(296-462): for BamHI CGCCACTAGTGGATCCATGTCTTCAACAATATCC for overlap CAGTCCTCGCAGAGTATGGAAATTATGAAG rev overlap CTTCATAATTTCCATACTCTGCGAGGACTG rev SalI TACCCTCGAGGTCGACTACAGTGCGTTTCCGCTT pSPYCE-Alb4(1-300)Alb3(314-462): for BamHI CGCCACTAGTGGATCCATGTCTTCAACAATATCC for overlap TCACAAGCAGTGACTCTTGTTTTCAAGTTT rev overlap AAACTTGAAAACAAGAGTCACTGCTTGTGA rev SalI TACCCTCGAGGTCGACTACAGTGCGTTTCCGCTT pSPYCE-Alb4(1-319)Alb3(336-462) entspricht pSPYCE-Alb4(1-302)Alb3(319-462): for BamHI CGCCACTAGTGGATCCATGTCTTCAACAATATCC for overlap CCTTCTGGTTTAAGTATTTACTGGCTCACA rev overlap TGTGAGCCAGTAAATACTTAAACCAGAAGG rev SalI TACCCTCGAGGTCGACTACAGTGCGTTTCCGCTT pSPYCE-Alb4(1-326)Alb3(343-462): for BamHI CGCCACTAGTGGATCCATGTCTTCAACAATATCC rev overlap TTGGGCGGTACTAAGTACGTT for overlap TATTGGCTGACCAACAACGTA rev SalI TACCCTCGAGGTCGACTACAGTGCGTTTCCGCTT pSPYCE-Alb3(1-342)Alb4(327-499): for BamHI CGCCACTAGTGGATCCATGGCGAGAGTTCTAGTC rev overlap TGCCGTGCTCAAAATATTATTTGTGAGCCA for overlap TGGCTCACAAATAATATTTTGAGCACGGCA rev SalI TACCCTCGAGGTCGACCCTCTTCTCTGTTTCATGAGA pSPYCE-Alb3(1-359): for BamHI CGCCACTAGTGGATCCATGGCGAGAGTTCTAGTC rev SalI TACCCTCGAGGTCGACCTTTGCACCACCTAGTTT pSPYCE-Alb3(1-366): for BamHI CGCCACTAGTGGATCCATGGCGAGAGTTCTAGTC rev SalI TACCCTCGAGGTCGACTGCGTTTTCATCCATATT pSPYCE-Alb3(1-415): for BamHI CGCCACTAGTGGATCCATGGCGAGAGTTCTAGTC rev SalI TACCCTCGAGGTCGACTTCTTCTACCAATTCAAC pSPYCE-Alb3(1-442): for BamHI CGCCACTAGTGGATCCATGGCGAGAGTTCTAGTC rev SalI TACCCTCGAGGTCGACGCTTGTTGTGCTCGAAGC pSPYCE-Alb3(F316T): for AATACGCTTCTTGTTACCAAGTTTCTTCCACTC rev GAGTGGAAGAAACTTGGTAACAAGAAGCGTATT
3 Material und Methoden 27
pSPYCE-Alb3(L314A): for CAGAAGAATACGCTTGCTGTTTTCAAGTTTCTT rev AAGAAACTTGAAAACAGCAAGCGTATTCTTCTG pSPYCE-Alb3(L313Q,L314A): for GCACAGAAGAATACGCAGGCTGTTTTCAAGTTTCTT rev AAGAAACTTGAAAACAGCCTGCGTATTCTTCTGTGC pSPYCE-Alb3(Δ314,315): for GCACAGAAGAATACGCTTTTCAAGTTTCTTCCACTC rev GAGTGGAAGAAACTTGAAAAGCGTATTCTTCTGTGC pSPYCE-Alb3(Δ314-317): for CAGAAGAATACGCTTTTTCTTCCACTCAT rev CATGAGTGGAAGAAAAAGCGTATTCTTCTG pSPYCE-Alb3(Δ374-388): for BamHI CGCCACTAGTGGATCCATGGCGAGAGTTCTAGTC rev overlap TTTTAATTGTCTAAATCCCGCACTTATTAT for overlap ATAATAAGTGCGGGATTTAGACAATTAAAA rev SalI TACCCTCGAGGTCGACTACAGTGCGTTTCCGCTT pSPYCE-Alb3(Δ389-415): for BamHI CGCCACTAGTGGATCCATGGCGAGAGTTCTAGTC rev overlap TGATTCAGACTGAGATCTTTCGCCAGCATC for overlap GATGCTGGCGAAAGATCTCAGTCTGAATCA rev SalI TACCCTCGAGGTCGACTACAGTGCGTTTCCGCTT pSPYCE-Alb3, pSPYNE-Alb3, pSPYCE-Alb4, pSPYNE-Alb4, pSPYCE-TS(rbcs)-cpSRP43, pSPYNE-TS(rbcs)-cpSRP43, pSPYCE-TS(rbcs)-cpSRP54, pSPYNE-TS(rbcs)-cpSRP54, pSPYCE-TS(rbcs)-cpFtsY, pSPYNE-TS(rbcs)-cpFtsY: (Laborbestand)
Nachfolgend gelistete Plasmide kamen bei den in vivo Interaktionsstudien im Hefe Split-
Ubiquitin System zum Einsatz.
pAMBV4-Alb3(Δ314-317): Template pSPYCE-Alb3(Δ314-317) for XbaI CTATCTAGACAAAAATGAGCTTAAACGAGATTCCTC rev Stu I ATGAGGCCTTTTACAGTGCGTTTCCGCTT pAMBV4-Alb3(Δ374-388): Template pSPYCE-Alb3(Δ374-388) for XbaI CTATCTAGACAAAAATGAGCTTAAACGAGATTCCTC rev Stu I ATGAGGCCTTTTACAGTGCGTTTCCGCTT pAMBV4-Alb3(Δ389-415): Template pSPYCE-Alb3(Δ389-415) for XbaI CTATCTAGACAAAAATGAGCTTAAACGAGATTCCTC rev Stu I ATGAGGCCTTTTACAGTGCGTTTCCGCTT pAMBV4-Alb3, pAMBV4-Alb4, pMBV-Alg5, pAlg5-NubI, pAlg5-NubG, pADSL-cpSRP43, pADSL-cpSRP54, pADSL-cpFtsY: (Laborbestand)
3 Material und Methoden 28
Des weiteren wurden folgende Sequenzierungsprimer verwendet:
pGEX4T3: for GGGCTGGCAAGCCACGTTTGGTG rev CCGGGAGCTGCATGTGTCAGAGG pETDuet1: for ATGCGTCCGGCGTAGAG rev GATTATGCGGCCGTGTACAA pSS: for GATTTAGGTGACACTATA rev TAATACGACTCACTATAGGG pIVEX1.3WG: for CGAAGTTGTAAAACGACGGCCAGT rev CAGGAAACAGCTATGAC pSPYCE: for AAGTTCATTTCATTTGGAGAG rev CTGCTTGTCAGCGTAATCTGG pSPYNE: for AAGTTCATTTCATTTGGAGAG rev CATAAGATCCTCCTCAGAAAT pAMBV4: for TTTCTGCACAATATTTCAAGC rev GTAAGGTGGACTCCTTCT
3 Material und Methoden 29
3.4.5 Mikroorganismen
3.4.5.1 Escherichia coli
Tabelle 3-6: Escherichia coli-Stämme
Stamm Genotyp Quelle
XL1 blue
recA1 endA1 gyrA96 thi-1 hsdR17, supE44 relA1 lac- [F´ pro AB lacIqZ∆M15 Tn10(tetr) ]
Abbildung 4-1: Alignment der Aminosäuresequenz von Alb3 und Alb4 Das Alignment zeigt einen Vergleich der Aminosäuresequenzen von Alb3 und Alb4. Dargestellt
sind die mittels ChloroP (http://www.cbs.dtu.dk/services/ChloroP/) vorhergesagten Transitpeptide
(TP) für das chloroplastidäre Targeting der Proteine (grün) sowie der maximal vorhergesagte
Bereich der fünf Transmembrandomänen (Aramemnon, http://aramemnon.botanik.uni-koeln.de/)
(grau hinterlegt). Die in blau gezeigten Sequenzen von Alb3 umfassen die Regionen der Alb3
Antigene nAlb3 (AS 56-108) und cAlb3 (AS 397-462), die zum Zweck der Antikörperherstellung
rekombinant überexprimiert wurden. Die in rot dargestellte Sequenz des Alb4 C-Terminus
beinhaltet den Bereich des Alb4 Antigens (AS 347-494).
4.1.1 Expression der Antigene GST-nAlb3, GST-cAlb3 und cAlb4-his
Für die Herstellung der Antigene von Alb3 wurden die entsprechenden Gensequenzen
des N-terminalen und C-terminalen Bereichs von Alb3 in den Expressionsvektor
pGEX4T3 kloniert und in den Überexpressionsstamm E. coli BL21(DE3) transformiert. Die
Antigene nAlb3 und cAlb3 wurden über diesen Vektor als GST-Fusionsproteine
exprimiert, da die errechnete Größe der Antigene mit 6 kDa (nAlb3) und 7 kDa (cAlb3) für
eine Immunisierung in Kaninchen nicht ausreichend war. Die Aufreinigung der Antigene
von Alb3 erfolgte über den angefügten GST-tag mittels Glutathion-Sepharose. Da für die
vorliegende Arbeit letztendlich lediglich der Antikörper gegen den C-Terminus von Alb3
4 Ergebnisse 65
eingesetzt wurde, ist die Funktionalität des Antikörpers gegen den N-Terminus nicht
abschließend getestet worden. Die Aufreinigung des Antigens GST-cAlb3 wurde über
eine SDS-PAGE mit anschließender Coomassie Färbung überprüft (Abbildung 4-2). Die
errechnete Größe des GST-Fusionsproteins beträgt 33 kDa und stimmt mit dem
tatsächlichen Laufverhalten bei 37 kDa gut überein. Eine weitere Proteinbande auf Höhe
von 32 kDa lässt auf ein Abbauprodukt des Proteins schließen. Das Problem des
Proteinabbaus bei rekombinant exprimiertem Alb3 Protein war bereits aus vorherigen
Versuchen im Labor bekannt. Die Gensequenz des Antigens von Alb4 (cAlb4) lag bereits
in dem Expressionsvektor pET29b in dem E. coli Überexpressionsstamm BL21(DE3) vor
(Gerdes et al., 2006). Nach der Überexpression erfolgte die Aufreinigung des Proteins
unter denaturierenden Bedingungen über den angefügten C-terminalen His-tag mittels Ni-
NTA Agarose. Anhand der Expressionskontrolle über SDS-PAGE mit anschließender
Coomassie Färbung zeigte sich ein zu der errechneten Größe des Proteins von 19 kDa
abweichendes Laufverhalten, das bei 26 kDa lag und bereits in Gerdes et al., 2006
beschrieben wurde (Abbildung 4-2). Nach erfolgter Aufreinigung wurden die Antigene
gegen PBS dialysiert und je 800 µg lyophilisiertes Protein zur Standardimmunisierung von
Kaninchen und Ratte an die Firma SeqLab (Göttingen) verschickt.
4.1.2 Test der Antikörper α-Alb3 und α-Alb4 gegen die Antigene
Nach Erhalt der Antiseren α-Alb3 (aus Kaninchen) und α-Alb4 (aus Ratte) wurden diese
zunächst gegen die Antigene getestet. Zu diesem Zweck wurde der GST-tag des cAlb3
Antigens durch Thrombin abgespalten und die erfolgreiche Abspaltung über eine SDS-
PAGE mit anschließender Coomassie Färbung überprüft (Abbildung 4-2). Nach dem
Thrombinverdau ist sowohl das 26 kDa große GST-Protein, als auch eine unterhalb von
17 kDa laufende Bande des abgespaltenen cAlb3 zu erkennen. Für den Antikörpertest
wurden das GST-cAlb3 Fusionsprotein, das gespaltene GST/cAlb3 Proteingemisch und
das cAlb4-his Protein über eine SDS-PAGE aufgetrennt und eine Immundetektion mit den
beiden Seren α-Alb3 (aus Kaninchen) und α-Alb4 (aus Ratte) durchgeführt. Mit dem
α-Alb3 Antikörper konnten das GST-cAlb3 Fusionsprotein und die durch Thrombin-
spaltung erhaltenen GST und cAlb3 Proteine detektiert werden. In der Spur, in der das
cAlb4 Antigen aufgetragen worden war, konnte kein Signal detektiert werden. Der Test
des α-Alb4 Antikörpers (aus Ratte) ergab, dass dieser das eigene cAlb4 Antigen und nicht
das GST-cAlb3 Fusionsprotein und die gespaltenen Proteine GST und cAlb3 detektierte.
Die Western Blots, die mit den Präimmunseren der beiden Antikörper behandelt wurden,
zeigten keine Signale. Diese Versuche belegen die Spezifität des α-Alb4 und des α-Alb3
Antikörpers im Hinblick auf die Detektion der verwendeten Antigene.
4 Ergebnisse 66
7255403324
17
kDa
α-Alb3 α-PI Alb3
α-PI Alb4α-Alb4
Coomassie
7255403324
17
kDa
Coomassie
**
*
*
*#
Abbildung 4-2: Test der Antikörper und Präimmunseren gegen die Antigene GST-cAlb3 und cAlb4-his
Für die Kontrolle der Aufreinigung der Antigene GST-cAlb3 und cAlb4-his wurden Aliquots der
Eluate über eine SDS-PAGE aufgetrennt und mit Coomassie angefärbt. Zur Überprüfung des
Thrombinverdaus wurde ein Aliquot des verdauten Proteingemisches GST/cAlb3 über SDS-PAGE
mit anschließender Coomassie Färbung analysiert. Für den Antikörpertest wurden die Antigene
GST-cAlb3 und cAlb4-his sowie das gespaltene GST/cAlb3 Proteingemisch über SDS-PAGE
aufgetrennt und eine Immundetektion mit den Antikörpern (α-Alb3 und α-Alb4) und den
entsprechenden Präimmunseren (α-PI Alb3 und α-PI Alb4) durchgeführt. (* Abbauprodukte des
GST-cAlb3 und cAlb4-his, # unspezifisches Signal)
4 Ergebnisse 67
4.1.3 Test der Antikörper α-Alb3 und α-Alb4 gegen die in vitro translatierten Volllängenproteine
Die Spezifität der α-Alb3 und α-Alb4 Antikörper sollte über den Test gegen die
Volllängenproteine in Form der in vitro Translationsprodukte des Alb3 und Alb4 bestätigt
werden (Abbildung 4-3). Dazu wurde das Alb3 und das Alb4 (mit Transitpeptid, pAlb4)
über die SP6-Polymerase in vitro transkribiert und anschließend mit Hilfe eines
Weizenkeimlysats in vitro translatiert. Die Translationskontrolle erfolgte über den Einsatz
von radioaktiv markiertem [35S]-Methionin, durch dessen Einbau die Proteine nach
Auftrennung mittels einer SDS-PAGE und anschließender Autoradiographie detektiert
werden konnten. Die Autoradiographie erbrachte in dem Translationsansatz des Alb3 ein
spezifisches Signal bei etwa 49 kDa und bei dem Ansatz des pAlb4 ein spezifisches
Signal bei etwa 60 kDa. Die Molekulargewichte dieser Signale entsprachen daher in etwa
den theoretischen Molekulargewichten von Alb3 (45 kDa) und dem pAlb4 inklusive des
Transitpeptids (55 kDa).
α-Alb3 α-Alb4
kDa
pAlb4
Alb3
72
55
43
Translations-kontrolle
Abbildung 4-3: Test der Antikörper gegen die in vitro translatierten Volllängenproteine
Für die Translationskontrolle wurden die radioaktiv markierten Translationsprodukte von Alb3 und
pAlb4 (TP [35S]-Alb3 und TP[35S]-pAlb4, 1 µl) über eine SDS-PAGE aufgetrennt und mittels
Autoradiographie nachgewiesen. Des Weiteren wurde die Spezifität der Antikörper α-Alb3 und
α-Alb4 mit nicht radioaktiv markierten Translationsprodukten (TP Alb3 und TP pAlb4, 5 µl) über
eine Immundetektion gezeigt.
Für den Antikörpertest wurden die Translationsansätze, in denen die Proteine nicht
radioaktiv markiert waren, über eine SDS-PAGE aufgetrennt und anschließend Western
Blot Analysen durchgeführt. Durch den Einsatz des α-Alb3 Antikörpers wurde das in vitro
translatierte Alb3 auf der Höhe von 49 kDa erkannt. Die Spezifität des α-Alb3 Antikörpers
4 Ergebnisse 68
wurde dadurch bestätigt, dass dieser Antikörper nicht das in vitro translatierte pAlb4
Volllängenprotein detektieren konnte. Zudem zeigte sich, dass der α-Alb4 Antikörper
ausschließlich das in vitro translatierte pAlb4 Volllängenprotein auf der Höhe von 60 kDa
und nicht das Translationsprodukt von Alb3 detektierte. Damit war es gelungen, die
Spezifität des α-Alb3 und α-Alb4 Antikörpers auch für das entsprechende
Volllängenprotein zu bestätigen.
4.1.4 Test der Antikörper α-Alb3 und α-Alb4 mit Thylakoidmembran-proteinen
Für den folgenden Antikörpertest wurden Thylakoidmembranproteine von Arabidopsis
thaliana über eine SDS-PAGE aufgetrennt und Western Blot Analysen durchgeführt
(Abbildung 4-4). Dabei wurde das Alb3 Protein durch den α-Alb3 Antikörper auf der Höhe
von 45 kDa nachgewiesen, während das auf der Höhe von 50 kDa zu erwartende Alb4
nicht detektiert wurde. Durch den Einsatz des α-Alb4 Antikörpers wurde ausschließlich
das Alb4 Protein auf der Höhe von 50 kDa erkannt. Die Western Blots, die mit den
Präimmunseren der beiden Antikörper behandelt wurden, zeigten keine Signale.
Insgesamt lässt sich festhalten, dass die in den Abschnitten 4.1.2 bis 4.1.4 beschriebenen
Versuche eindeutig die Spezifität der beiden Antikörper gegen Alb3 und Alb4 belegen.
Damit war eine wichtige Voraussetzung für weitere biochemische Analysen von Alb3 und
Alb4 erfüllt.
α-Alb4
kDaTM TMTM TM
α-PI Alb4α-PI Alb3α-Alb3
72
55
43
95
Abbildung 4-4: Test der Antikörper gegen Thylakoidmembranproteine Zur Überprüfung der Spezifität der Antikörper wurden Thylakoidmembranproteine (äquivalent zu
5 µg Chlorophyll) über eine SDS-PAGE aufgetrennt und ein immunologischer Nachweis mit den
Antikörpern α-Alb3 und α-Alb4 durchgeführt. Weitere Western Blot Analysen dienten der Kontrolle
auf Reaktion der Präimmunseren (α-PI Alb3 und α-PI Alb4) mit den Thylakoidmembranproteinen.
4 Ergebnisse 69
4.2 Analyse der Interaktionen von Alb3 und Alb4 mit den Proteinen des cpSRP-Transportwegs
Bisherige Studien zeigten, dass der Transport der Lichtsammelkomplexproteine (LHCP)
durch das Stroma zur Thylakoidmembran über den cpSRP-Transportweg erfolgt. Des
Weiteren stellt das integrale Membranprotein Alb3 die Translokase für die Integration der
LHCPs in die Membran dar. Ein entscheidender Aspekt innerhalb dieses Transportwegs,
der sich auf den docking-Prozess des Transitkomplexes an die Thylakoidmembran
bezieht, ist allerdings bisher noch ungeklärt. In dem folgenden Teil dieser Arbeit sollte
geklärt werden, ob der docking-Prozess des Transitkomplexes möglicherweise durch eine
direkte Interaktion der Proteine des cpSRP-Wegs mit der Translokase Alb3 vermittelt wird.
Außerdem sollte über die Proteininteraktionsstudien untersucht werden, ob das zur
Translokase Alb3 homologe Protein Alb4 eine Funktion innerhalb des cpSRP-
Transportwegs ausübt.
4.2.1 Interaktionen von Alb3 und Alb4 mit den Proteinen des cpSRP-Transportwegs im Hefe Split-Ubiquitin System
Um die Interaktion der beiden Membranproteine Alb3 und Alb4 mit den Proteinen des
cpSRP-Wegs cpSRP43, cpSRP54 und cpFtsY zu analysieren, wurde zunächst das Hefe
Split-Ubiquitin System eingesetzt. Das Split-Ubiquitin System ist ein in vivo Verfahren, bei
dem die Interaktion von Membranproteinen getestet werden kann. Dabei ist das Ubiquitin
in ein N-terminales Nub-Fragment und ein C-terminales Cub-Fragment gespalten. Das
N-terminale Fragment des Ubiquitins (NubI) besitzt eine hohe Affinität zum C-terminalen
Bereich des Ubiquitins (Cub). Bei räumlicher Nähe eines Fusionsproteins, bestehend aus
dem Cub-Fragment und dem zu analysierenden Protein (Alb3 bzw. Alb4), und dem NubI-
Fragment des Kontrollproteins Alg5-NubI kommt es zur Rekonstitution des Ubiquitins und
damit zur Abspaltung des Reporters. Dieser Reporter ist an das Cub-Protein angefügt und
besteht aus einer DNA-bindenden Domäne (LexA-Protein) und einer
transkriptionsaktivierenden Domäne (Virionenprotein 16). Die Lokalisation des Reporters
im Cytosol kann durch eine erfolgte Aktivierung der Reportergene nachgewiesen werden.
Zur Kontrolle auf Transaktivierung diente das Protein Alg5-NubG, das aufgrund einer
Punktmutation im Nub-Fragment (NubG) die Affinität zwischen den Ubiquitin-Hälften
verringert. Mit Hilfe des Split-Ubiquitin Systems wurden bereits erfolgreiche
Untersuchungen des Alb3 mit potentiellen Bindungspartnern durchgeführt (Pasch et al.,
2005). Das Cub-Protein wird mit dem angefügten Reporter von dem pAMBV4-Vektor und
das NubG-Protein durch den Vektor pADSL-Nx kodiert. Für den Interaktionstest wurden
die Gensequenzen von Alb3 und Alb4 in den pAMBV4-Vektor und die Gensequenzen von
4 Ergebnisse 70
cpSRP43, cpSRP54 und cpFtsY in den pADSL-Vektor kloniert. Die Kontrolle der
Cotransformation der bait- und prey-Plasmide in die Hefezellen verlief in allen Ansätzen
positiv. Die Transformationsraten konnten durch das Wachstum der Hefekolonien auf dem
–LT-Medium, dem die Aminosäuren Leucin und Tryptophan fehlten, erkannt werden
(Daten nicht gezeigt). Der Einsatz der Konstrukte pAMBV4-Alb3 bzw. pAMBV4-Alb4 mit
dem Kontrollplasmid (kodiert für Alg5-NubI) wies durch ein deutliches Wachstum der
cotransformierten Hefezellen auf dem –LTH-Medium auf eine erfolgreiche Rekonstitution
des Ubiquitins hin. Zur Kontrolle auf Transaktivierung des Alb3 und Alb4 wurden die
Plasmide pAMBV4-Alb3 bzw. pAMBV4-Alb4 mit dem Kontrollplasmid (kodiert für Alg5-
NubG) cotransformiert. Ein Wachstum der transformierten Hefezellen auf –LTH-Medium
wies auf eine schwache Eigenaktivierung hin (Daten nicht gezeigt). Der Zusatz von
15 mM 3-Aminotriazol (3-AT), einem kompetitiven Inhibitor des His3-Genproduktes,
verringerte die Eigenaktivierung jedoch so weit, dass kein Wachstum beim Einsatz des
Kontrollplasmids auf –LTH-Medium mehr möglich war.
Für die Interaktionsanalysen (Tabelle 4-1) wurden die Konstrukte pAMBV4-Alb3 bzw.
pAMBV4-Alb4 sowie die Kontrolle pMBV4-Alg5 mit den Konstrukten der potentiellen
Bindungspartner (pADSL-cpSRP43, pADSL-cpSRP54 und pADSL-cpFtsY) in die
Hefezellen cotransformiert. Interessanterweise deuteten die Ergebnisse auf eine
Interaktion des Alb3 mit cpSRP43, während eine Interaktion mit cpSRP54 oder cpFtsY im
Split-Ubiquitin System nicht nachgewiesen wurde. Um auszuschließen, dass die
negativen Ergebnisse auf eine fehlende Expression des cpSRP54 oder cpFtsY
zurückzuführen sind, wurde die korrekte Expression der Proteine in den Hefezellen
nachgewiesen (Daten nicht gezeigt). Das Alb4 zeigte mit keinem der getesteten Proteine
eine Interaktion im Hefe Split-Ubiquitin System. Von den Hefekolonien, die auf dem –LTH-
Medium gewachsen sind, wurde ein β-Galaktosidase Filtertest durchgeführt. Bei allen
Transformationen, bei denen ein Wachstum der Hefezellen auf diesem Selektionsmedium
zu verzeichnen war, konnte auch eine Blaufärbung im Filtertest gezeigt werden (Tabelle
4-1). Diese Ergebnisse deuten darauf hin, dass das cpSRP43 in der Lage ist direkt mit
Alb3 zu interagieren und dass diese Interaktion möglicherweise den docking-Prozess des
Transitkomplexes an die Thylakoidmembran über eine Interaktion mit der Translokase
Alb3 vermittelt.
4 Ergebnisse 71
Tabelle 4-1: Alb3 interagiert mit cpSRP43 im Split-Ubiquitin System Dargestellt sind die Interaktionstests von Alb3 und Alb4 mit den löslichen Proteinen des cpSRP-
Wegs im Hefe Split-Ubiquitin System. Der Hefe Stamm DSY-1 wurde mit den Plasmiden pAMBV4-
Alb3, pAMBV4-Alb4, pMBV4-Alg5 und den angegebenen NubG-Fusionsproteinen des cpSRP-
Wegs cotransformiert und die Aktivierung der Reportergene ausgewertet. Durch das Wachstum
der cotransformierten Hefezellen auf Minimalmedium (-his) (++ = deutliches Wachstum,
+ = geringes Wachstum, - = kein Wachstum) und über die Bestimmung der Enzymaktivität der
β-Galaktosidase (β-gal), die durch eine Blaufärbung der Hefekolonien im β-Galaktosidase Filtertest
getestet wurde (+ = Blaufärbung, - = keine Blaufärbung), konnten Aussagen über die Interaktion
der Proteine gemacht werden. Die Plasmide pADSL-Alg5-NubI und pADSL-Alg5-NubG wurden als
Positiv- bzw. Negativkontrollen eingesetzt.
pAMBV4-Alb3 pAMBV4-Alb4 pMBV4-Alg5
pADSL-Nx -his β-gal. -his β-gal. -his β-gal
Alg5-NubI ++ + ++ + + +
Alg5-NubG - - - - - -
cpSRP43 ++ + - - - -
cpSRP54 - - - - - -
cpFtsY - - - - - -
4.2.2 Proteininteraktionen von Alb3 und Alb4 mit den Proteinen des cpSRP-Transportwegs im Split-YFP System
Um die gezeigte in vivo Interaktion von Alb3 mit cpSRP43 zu bestätigen, sollten die im
Split-Ubiquitin System getesteten Proteine im Split-YFP System in Arabidopsis thaliana
Protoplasten eingesetzt werden. Dieses ist ein weiteres in vivo Verfahren, um mit Hilfe
der bimolekularen Fluoreszenzkomplementation die Interaktion von Proteinen in ihrer
natürlichen Umgebung zu testen. Dazu ist das YFP in zwei Hälften gespalten. Das N-
terminale nYFP wird durch den Vektor pSPYNE (split YFP N-terminal fragment
expression) und das C-terminale cYFP durch den Vektor pSPYCE (split YFP
C-terminal fragment expression) kodiert. Für die Interaktionstests wurden die
Membranproteine Alb3 und Alb4 N-terminal an die YFP-Fragmente fusioniert, so dass die
YFP-Hälften nach der Integration der Proteine in die Thylakoidmembran auf der Stroma
gelegenen Seite lokalisiert sind. Damit war gewährleistet, dass die YFP-Fragmente der
Membranproteine mit den entsprechenden YFP-Hälften der möglichen stromalen
Interaktionspartner cpSRP43, cpSRP54 und cpFtsY in räumliche Nähe geraten konnten.
Der Transport der Proteine (Alb3, Alb4 und cpFtsY) aus dem Cytosol in den Chloroplasten
4 Ergebnisse 72
wurde über die jeweilige chloroplastidäre Transitsequenz der Proteine vermittelt. Lagen
die Gensequenzen der Proteine ohne eigene Transitsequenz vor (cpSRP43 und
cpSRP54), wurde die Transitsequenz der kleinen Untereinheit der Rubisco (TS-rbcs) vor
das jeweilige Konstrukt kloniert, um den chloroplastidären Import zu erreichen. Für die
Analyse der Interaktion wurde die Gensequenz des einen Interaktionspartners in den
pSPYCE-Vektor und die des anderen Partners in den pSPYNE-Vektor kloniert und diese
Konstrukte in Arabidopsis thaliana Protoplasten cotransformiert. Bei einer Interaktion
zweier YFP-Fusionsproteine rekonstituieren sich die beiden nicht-fluoreszierenden YFP-
Hälften zu dem fluoreszierenden YFP, dessen Fluoreszenz an einem Laserscanning
Mikroskop (LSM 510 Meta) visualisiert und dokumentiert werden konnte.
Zunächst sollte getestet werden, ob die bereits bekannte Interaktion von cpSRP43 und
cpSRP54 mit diesem System gemessen werden kann. In Abbildung 4-5 ist gezeigt, dass
cpSRP43-nYFP mit dem cpSRP54-cYFP in Arabidopsis thaliana Protoplasten
interagierte, wodurch die Fluoreszenz des YFP rekonstituiert werden konnte. Die
Überlagerung der YFP-Fluoreszenz mit der Autofluoreszenz des Chlorophylls zeigte
zudem die Interaktion der beiden Proteine innerhalb der Chloroplasten. Es zeigte sich
ebenfalls eine Interaktion von Alb3-cYFP mit cpSRP43-nYFP und Alb3-nYFP mit
cpSRP43-cYFP in vivo in Arabidopsis Protoplasten, wodurch das Ergebnis der Interaktion
der beiden Proteine im Hefe Split-Ubiquitin System bestätigt werden konnte (Abbildung
4-5, Tabelle 4-3). Als Kontrolle wurden die Vektoren pSPYNE-TS(rbcs) und pSPYCE-
TS(rbcs) eingesetzt und mit den einzelnen Fusionskonstrukten cotransformiert. Bei diesen
Konstrukten ist die Transitsequenz der kleinen Untereinheit der Rubisco (TS-rbcs) vor das
jeweilige YFP-Fragment fusioniert, wodurch der Import der einzelnen YFP-Hälften ohne
weiteres Fusionsprotein in die Chloroplasten garantiert ist. Die Kontrollen zeigten, dass
weder das Fusionsprotein Alb3-cYFP noch das cpSRP43-nYFP in der Lage waren die
YFP-Fluoreszenz mit dem jeweiligen nYFP- oder cYFP-Fragment zu rekonstituieren
(Abbildung 4-5, Tabelle 4-2). Diese Ergebnisse deuteten auf eine direkte Interaktion
zwischen Alb3 und cpSRP43, da eine unspezifische und zufällige Rekonstitution des
YFPs auszuschließen war.
Des Weiteren sollten die Proteine cpSRP54 und cpFtsY auf Interaktion mit Alb3 getestet
werden. Es konnte keine YFP-Fluoreszenz und damit keine Interaktion von Alb3 mit
cpSRP54 oder cpFtsY im Split-YFP System erkannt werden. Zudem zeigte sich, dass
Alb4 im Vergleich zu Alb3 nicht in der Lage war mit cpSRP43 zu interagieren (Tabelle
4-2). Alle Proteine wurden sowohl als Fusionsproteine mit der N-terminalen Hälfte, als
auch als Fusionsproteine mit der C-terminalen Hälfte des YFP getestet. Dazu lagen die
Gensequenzen für diese Proteine im pSPYCE- und pSPYNE-Vektor vor (Tabelle 4-2). In
allen Kontrollexperimenten konnte ebenfalls keine YFP-Rekonstitution beobachtet werden
(Tabelle 4-3).
4 Ergebnisse 73
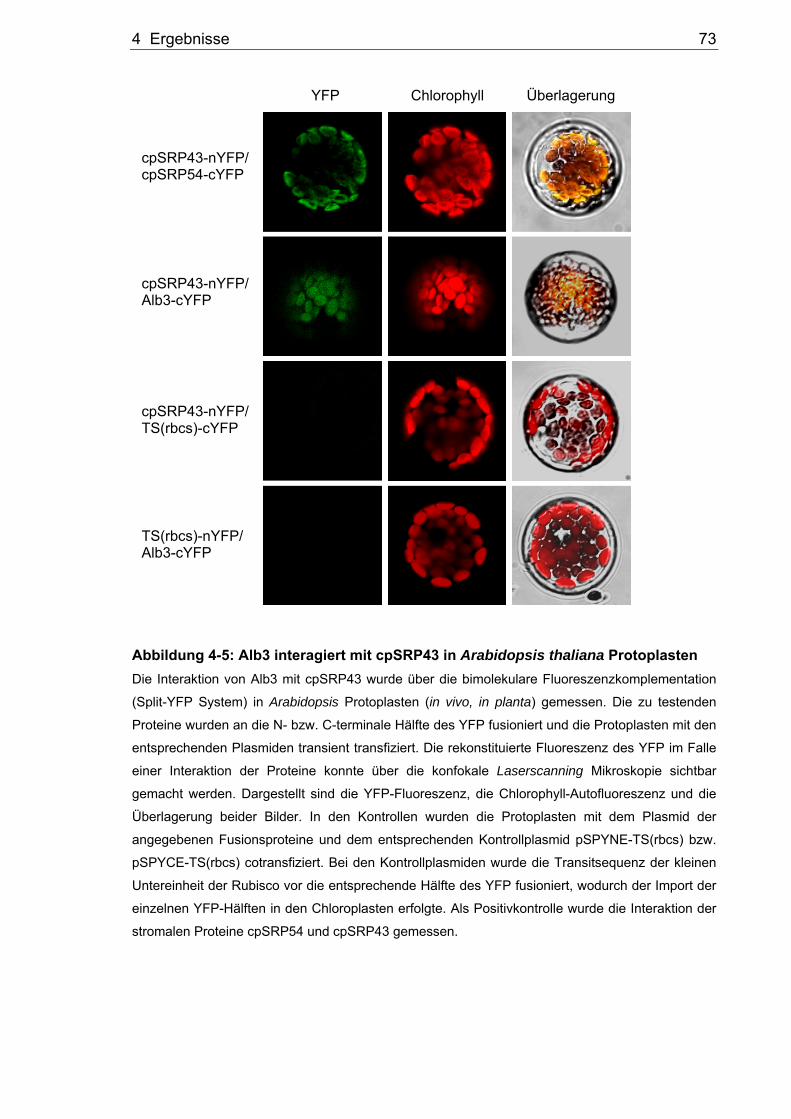
Abbildung 4-5: Alb3 interagiert mit cpSRP43 in Arabidopsis thaliana Protoplasten Die Interaktion von Alb3 mit cpSRP43 wurde über die bimolekulare Fluoreszenzkomplementation
(Split-YFP System) in Arabidopsis Protoplasten (in vivo, in planta) gemessen. Die zu testenden
Proteine wurden an die N- bzw. C-terminale Hälfte des YFP fusioniert und die Protoplasten mit den
entsprechenden Plasmiden transient transfiziert. Die rekonstituierte Fluoreszenz des YFP im Falle
einer Interaktion der Proteine konnte über die konfokale Laserscanning Mikroskopie sichtbar
gemacht werden. Dargestellt sind die YFP-Fluoreszenz, die Chlorophyll-Autofluoreszenz und die
Überlagerung beider Bilder. In den Kontrollen wurden die Protoplasten mit dem Plasmid der
angegebenen Fusionsproteine und dem entsprechenden Kontrollplasmid pSPYNE-TS(rbcs) bzw.
pSPYCE-TS(rbcs) cotransfiziert. Bei den Kontrollplasmiden wurde die Transitsequenz der kleinen
Untereinheit der Rubisco vor die entsprechende Hälfte des YFP fusioniert, wodurch der Import der
einzelnen YFP-Hälften in den Chloroplasten erfolgte. Als Positivkontrolle wurde die Interaktion der
stromalen Proteine cpSRP54 und cpSRP43 gemessen.
YFP Chlorophyll Überlagerung
cpSRP43-nYFP/ cpSRP54-cYFP
cpSRP43-nYFP/ Alb3-cYFP
cpSRP43-nYFP/ TS(rbcs)-cYFP
TS(rbcs)-nYFP/ Alb3-cYFP
4 Ergebnisse 74
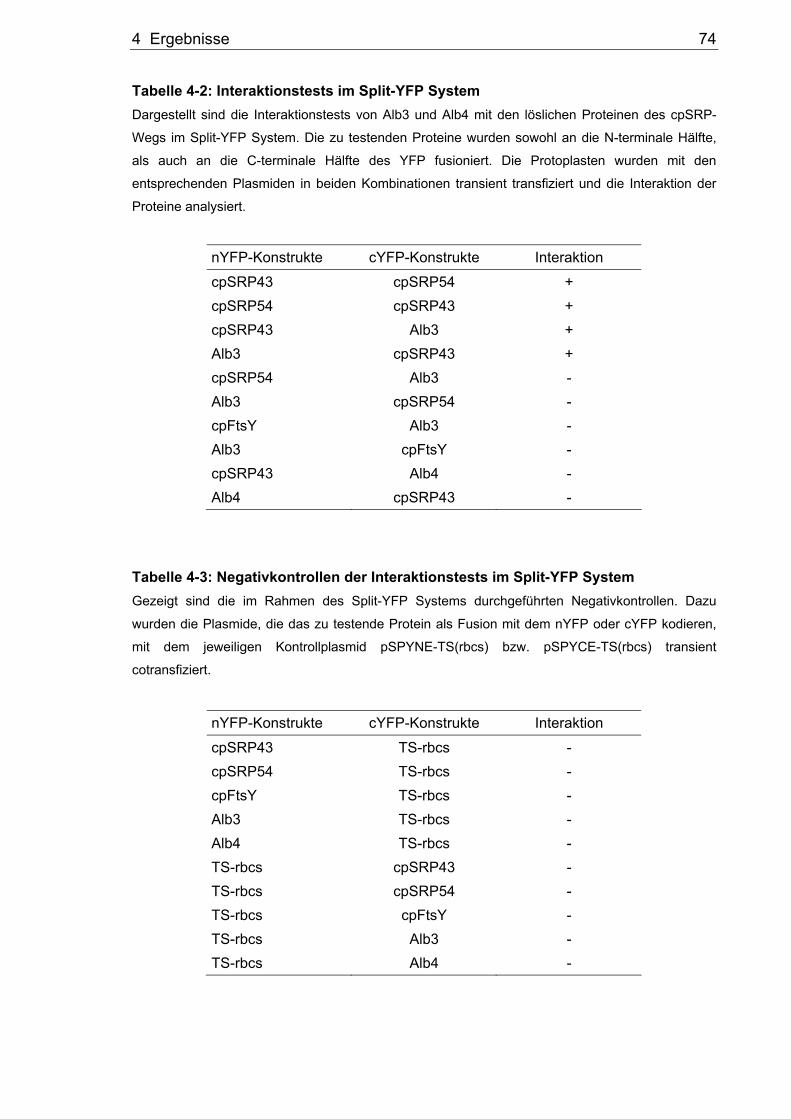
Tabelle 4-2: Interaktionstests im Split-YFP System Dargestellt sind die Interaktionstests von Alb3 und Alb4 mit den löslichen Proteinen des cpSRP-
Wegs im Split-YFP System. Die zu testenden Proteine wurden sowohl an die N-terminale Hälfte,
als auch an die C-terminale Hälfte des YFP fusioniert. Die Protoplasten wurden mit den
entsprechenden Plasmiden in beiden Kombinationen transient transfiziert und die Interaktion der
Tabelle 4-3: Negativkontrollen der Interaktionstests im Split-YFP System Gezeigt sind die im Rahmen des Split-YFP Systems durchgeführten Negativkontrollen. Dazu
wurden die Plasmide, die das zu testende Protein als Fusion mit dem nYFP oder cYFP kodieren,
mit dem jeweiligen Kontrollplasmid pSPYNE-TS(rbcs) bzw. pSPYCE-TS(rbcs) transient
Abbildung 4-10: Peptide Scanning von Alb3 Peptiden mit rekombinantem cpSRP43 Für die Methode des Peptide Scanning wurde zunächst das cpSRP43 rekombinant überexprimiert
und über den angefügten N-terminalen His-tag aufgereinigt. Ein Aliquot des Eluats His-cpSRP43
(His-43) wurde über eine SDS-PAGE aufgetrennt und anschließend mit Coomassie gefärbt, um die
erfolgreiche Aufreinigung zu überprüfen (A). Die Interaktion von rekombinantem His-cpSRP43 und
den Alb3 Peptiden wurde über eine Pepscan Methode (PepSpots, JPT Peptide Technologies,
Berlin) analysiert. Die dargestellte Sequenz von Alb3 zeigt die Aminosäuren, die für die PepSpot
Membran eingesetzt wurden (blau) (B). Der Bereich umfasst den 2. lumenalen loop, die
5. Transmembrandomäne (grau: Consensus Sequenz, hellgrau: zusätzliche Aminosäuren der
maximalen Vorhersage) sowie den C-Terminus von Alb3. Die Membran enthält 51 Peptide mit
jeweils 15 Aminosäuren Länge, die jeweils um drei Aminosäuren versetzt aufgetragen waren. Das
gebundene, rekombinante His-cpSRP43 wurde über einen Western Blot mit Hilfe eines Antikörpers
gegen das cpSRP43 (α-chaos) detektiert (C). Die interagierenden Peptide 3-6 umfassen die
Aminosäuren 305-328 und das Peptid 26 die Aminosäuren 374-388 von Alb3 und sind als
Bindestellen in der Alb3 Sequenz unterstrichen dargestellt (B). Die Analyse der PepSpots 3-6
erbrachte ein gemeinsames Bindemotiv der Aminosäuren 314-319 (LVFKFL) und ist in rot
hervorgehoben (D).
4 Ergebnisse 82
4.2.5 Bestätigung der mittels Peptide Scanning gezeigten Binderegionen im Split-YFP System
Die Auswertung des Peptide Scanning bestätigte die bereits über die Fusionskonstrukte
von Alb4 und Alb3 im Split-YFP gezeigte Bindestelle innerhalb der
5. Transmembrandomäne von Alb3. Aus diesem Grund wurden gezielt Mutationen und
Deletionen in diesem Bereich des Alb3 erzeugt und diese Proteine auf Interaktion mit
cpSRP43 im Split-YFP System getestet. Auf Grundlage eines Alignments der
Proteinsequenzen von Alb3 und Alb4 wurden die Aminosäuren von Alb3 innerhalb des
putativen Bindemotivs (LVFKFL) zu den entsprechenden Aminosäuren der Alb4 Sequenz
Abbildung 4-11: Alignment von Alb3 und Alb4 im Bereich des Bindemotivs Das Alignment von Alb3 und Alb4 zeigt die Aminosäuren (rot), die bei den Mutationskonstrukten
Alb3(F316T), Alb3(L314A) und Alb3(L313Q,L314A) gegeneinander ausgetauscht worden sind.
Während das mutierte Protein Alb3(F316T) noch in der Lage war durch Interaktion mit
cpSRP43 die YFP-Fluoreszenz zu rekonstituieren, konnten die Proteine Alb3(L314A) und
Alb3(L313Q,L314A) nicht mehr mit dem cpSRP43 interagieren (Tabelle 4-4). Die
Deletionskonstrukte Alb3(Δ314,315) und Alb3(Δ314-317) zeigten ebenfalls keine
Interaktion mit dem cpSRP43, was auf eine entscheidende Bedeutung dieser Bindestelle
innerhalb der 5. Transmembrandomäne deutet (Tabelle 4-4). Die Analyse des Peptide
Scanning ergab weiterhin, dass die Aminosäuren 374-388 eine Bindestelle für das
cpSRP43 auf dem C-Terminus von Alb3 bilden. Um dieses zu bestätigen wurden die
Aminosäuren 374-388 mittels overlap-PCR in dem Alb3 Protein deletiert und die
Interaktion mit dem cpSRP43 ebenfalls über die bimolekulare Fluoreszenzkomplemetation
getestet. Die Deletion der Aminosäuren 374-388 hatte zur Folge, dass das Alb3(Δ374-
388) nicht mehr mit cpSRP43 interagieren konnte (Tabelle 4-4). Als Kontrolle wurde mit
den Aminosäuren 389-415 ein weiterer Bereich des C-Terminus von Alb3 deletiert. Das
Protein Alb3(Δ389-415) konnte im Vergleich zu dem Alb3(Δ374-388) noch mit dem
cpSRP43 interagieren (Tabelle 4-4). Die Aminosäuren 389-415 haben daher keinen
Einfluss auf die Bindung des cpSRP43. In weiteren Versuchen wurden C-terminal
verkürzte Alb3 Proteine im Split-YFP System getestet. Man konnte feststellen, dass die
4 Ergebnisse 83
Proteine Alb3(1-359) und Alb3(1-366) nicht mehr mit dem cpSRP43 interagieren konnten,
währenddessen die Proteine Alb3(1-415) und Alb3(1-442) eine Interaktion zeigten
(Tabelle 4-4). Damit führte die C-terminale Verkürzung des Proteins von 415 auf 366
Aminosäuren zum Ausbleiben der Bindung von cpSRP43. Da sich innerhalb dieses
deletierten Bereiches die gezeigte Bindestelle der Aminosäuren 374-388 befindet,
unterstützen auch diese Ergebnisse die Präsenz dieses Bindemotivs im C-Terminus von
Alb3.
Tabelle 4-4: Interaktion der Alb3 Deletions- und Mutationsproteine mit cpSRP43-nYFP in Arabidopsis Protoplasten
Die mittels overlap-PCR und Mutagenese-PCR erzeugten Deletions- und Mutationsproteine von
Alb3 wurden im Split-YFP System auf Interaktion mit cpSRP43 getestet. Die Alb3 Konstrukte
wurden an das cYFP und das cpSRP43 an das nYFP fusioniert. Arabidopsis Protoplasten wurden
mit den angegebenen Konstrukten transient transfiziert und die Proteine auf Interaktion analysiert.
Um eine Assoziation von Alb4 mit der ATP-Synthase zu verifizieren, wurden die
Proteinkomplexe solubilisierter Thylakoidmembranen über eine Gelfiltrations-
chromatographie fraktioniert. Die Präsenz der Proteine Alb4, Alb3, cpSecY, CF1β, CF0II
und CF0III in den einzelnen Fraktionen wurde mittels Immundetektion über spezifische
Antikörper ausgewertet (Abbildung 4-21). Die Western Blot Analysen zeigten, dass der
überwiegende Teil von Alb4 mit den ATP-Synthase Untereinheiten CF1β, CF0II und CF0III
bei einem Molekulargewicht von 360-450 kDa von der Gelfiltrationssäule coeluierte.
Weiterhin konnte aus vorangegangenen Arbeiten (Klostermann et al., 2002) bestätigt
werden, dass die Alb3 und cpSecY beinhaltenden Proteinkomplexe mit einer Größe von
etwa 150-200 kDa von der Gelfiltrationsmatrix eluiert wurden. Diese Fraktionen
überlappten nur geringfügig mit den Alb4 Eluaten. Somit konnte gezeigt werden, dass
Alb4 mit der ATP-Synthase cofraktioniert und nur geringfügig mit Alb3 oder cpSecY.
~150 kDa ~15 kDaTM
~360 kDa
Alb3
cpSecY
Alb4
CF1β
~3600 kDa
CF0III
CF0II
Abbildung 4-21: Alb4 cofraktioniert mit den ATP-Synthase Untereinheiten Thylakoidmembranen (500 µg Chlorophyll) wurden mit 1,5 % Dodecyl-β-D-maltosid solubilisiert
und die Proteinkomplexe über eine Gelfiltrationssäule in 1 ml Fraktionen eluiert. Aliquots der
Fraktionen und solubilisierter Thylakoidmembranen wurden über eine SDS-PAGE aufgetrennt und
mittels spezifischer Antikörper gegen Alb4, Alb3, cpSecY, CF1β, CF0II und CF0III in einer
Anhand von Coimmunpräzipitationsexperimenten sollte zunächst bestätigt werden, dass
Alb4 mit der ATP-Synthase assoziiert vorliegt und keine Komponente der Alb3 und
cpSecY beinhaltenden Proteinkomplexe ist. Dazu wurden solubilisierte Thylakoid-
membranen mit den an Protein-A-Sepharose gekoppelten Antikörpern α-Alb3, α-Alb4 und
α-cpSecY inkubiert. Die Präzipitate wurden über eine SDS-PAGE aufgetrennt und mittels
Immundetektion analysiert (Abbildung 4-22). Da die Präzipitate nicht vollständig frei von
den Antikörper IgGs waren, musste für die Präzipitation des Alb4 der α-Alb4 Antikörper
aus Kaninchen und für die Detektion des Alb4 der in dieser Arbeit bereits beschriebene
α-Alb4 Antikörper aus Ratte verwendet werden. Dieses war erforderlich, da die IgGs mit
einer Größe von etwa 55 kDa auf der gleichen Höhe wie das Alb4 liefen. Der Nachweis
mit dem Antikörper aus Kaninchen wäre somit nicht möglich gewesen, da die IgGs das
Alb4 bei einer Immundetektion überdecken würden. Die Herstellung eines spezifischen
α-Alb4 Antikörpers aus Ratte (oder einem anderen Organismus) war für die Methode der
Coimmunpräzipitation von entscheidender Bedeutung und Notwendigkeit.
TM IP
cpSecY
Alb3
Alb4
Abbildung 4-22: Coimmunpräzipitation von Alb3, Alb4 und cpSecY Thylakoidmembranen (500 µg Chlorophyll) wurden mit 1,5 % Digitonin solubilisiert und mit den an
Protein-A-Sepharose gekoppelten α-Alb3, α-Alb4 (Kaninchen) und α-cpSecY Antikörpern
immunpräzipitiert. Aliquots der Coimmunpräzipitate und solubilisierter Thylakoidmembranen
wurden über eine SDS-PAGE aufgetrennt und mittels Immundetektion analysiert. Der Nachweis
des Alb4 in den Präzipitaten erfolgte mit einem α-Alb4 Antikörper aus Ratte zur Vermeidung der
Detektion der Kaninchen IgGs auf Höhe des Alb4 Proteins. (IP: Immunpräzipitation, TM:
solubilisierte Thylakoidmembranen)
4 Ergebnisse 102
Die Western Blot Analysen zeigten, dass bei Verwendung des α-Alb3 Antikörpers das
Alb3 zusammen mit cpSecY copräzipitiert werden konnte. Gleiches gilt bei Einsatz des α-
cpSecY Antikörpers, wodurch die Coimmunpräzipitation der Proteine Alb3 und cpSecY
gezeigt werden konnte. Dieses Ergebnis ist eine Bestätigung der bereits in Klostermann
et al., 2002 veröffentlichten Daten. Weiterhin zeigte sich, dass weder Alb3 noch cpSecY
mit dem Alb4 unter Verwendung des α-Alb4 Antikörpers coimmunpräzipitiert werden
konnte. Auch die Präzipitate des α-Alb3 Antikörpers und α-cpSecY Antikörpers
beinhalteten kein Alb4 Protein. Dieses unterstützt die Aussage, dass die Proteine Alb3
und Alb4 in verschiedenen Thylakoidmembrankomplexen assoziiert sind. Weiterhin
musste allerdings festgestellt werden, dass die Analyse der Alb4 Präzipitate keine
Coimmunpräzipitation von Alb4 mit den ATP-Synthase Untereinheiten zeigte. Die
Assoziation von Alb4 mit der ATP-Synthase konnte daher mittels Coimmunpräzipitation
nicht bestätigt werden.
5 Diskussion 103
5. Diskussion
Die Mitglieder der Oxa1/YidC/Alb3-Proteinfamilie haben eine essentielle Bedeutung für
der Insertion und Assemblierung von Membranproteinen in Mitochondrien, Bakterien und
Chloroplasten (Kuhn et al., 2003; Xie und Dalbey, 2008). In der vorliegenden Arbeit sollte
die Rolle der chloroplastidären Homologe der Oxa1/YidC/Alb3-Proteinfamilie (Alb3 und
Alb4) bei der Biogenese der Thylakoidmembran von Arabidopsis thaliana analysiert
werden. Das integrale Thylakoidmembranprotein Alb3 ist für die posttranslationale
Insertion der Lichtsammelkomplexproteine (LHCPs) verantwortlich (Sundberg et al., 1997;
Moore et al., 2000). Zudem ist Alb3 mit der cpSec-Translokase assoziiert und scheint
daher an der cotranslationalen Insertion von plastidär kodierten Membranproteinen
beteiligt zu sein (Klostermann et al., 2002). Das Alb4 ist ebenfalls in der
Thylakoidmembran lokalisiert und hat einen Einfluss auf die Biogenese der Chloroplasten
(Gerdes et al., 2006). Die genaue Funktion von Alb4 ist jedoch noch nicht geklärt.
5.1 Die cpSRP-Untereinheit cpSRP43 interagiert direkt mit dem integralen Membranprotein Alb3
Bisherige Studien zum posttranslationalen cpSRP-Transportweg haben gezeigt, dass das
cpSRP43, cpSRP54, cpFtsY, GTP und die Translokase Alb3 für die Integration der
LHCPs in die Thylakoidmembran notwendig sind (Tu et al., 1999; Yuan et al., 2002). Das
cpSRP54 und der cpSRP-Rezeptor cpFtsY sind zusammen in der Lage mit dem
integralen Membranprotein Alb3 einen Komplex an der Thylakoidmembran zu bilden
(Moore et al., 2003). Moore et al., 2003 zeigten weiterhin, dass weder das Substrat LHCP,
noch das cpSRP43 für die Ausbildung dieses Komplexes nötig waren. Eine direkte
Interaktion zwischen cpSRP43 und Alb3 wurde durch die angewendeten Crosslinking
Studien nicht nachgewiesen. Damit wurde vermutet, dass das cpSRP43 für den docking-
Prozess des Transitkomplexes an die Thylakoidmembran keinen entscheidenden Einfluss
hat.
Einige Fragen, wie z. B. der docking-Prozess des Transitkomplexes an die
Thylakoidmembran, die Freisetzung und Insertion der LHCPs sowie die Regulation des
cpSRP-Transportwegs sind noch nicht ausreichend erforscht. In der vorliegenden Arbeit
sollte zunächst untersucht werden, ob der docking-Prozess des Transitkomplexes an die
Thylakoidmembran möglicherweise durch eine direkte Interaktion der löslichen Proteine
des cpSRP-Wegs mit der Translokase Alb3 vermittelt wird.
Die Ergebnisse der in vivo Interaktionsstudien über das Hefe Split-Ubiquitin System und
die bimolekulare Fluoreszenzkomplementation (Split-YFP System) zeigten, im Vergleich
zu den zuvor beschriebenen Beobachtungen aus Moore et al., 2003, eine direkte
5 Diskussion 104
Interaktion von cpSRP43 mit Alb3 (Kapitel 4.2.1 und 4.2.2). Dieses war sowohl in der
Hefe, als auch in der natürlichen Umgebung der Arabidopsis thaliana Protoplasten der
Fall. Es wurde ausschließlich eine Interaktion des cpSRP43 mit dem Alb3 nachgewiesen,
während das cpSRP54 und cpFtsY mit Alb3 keine Interaktion zeigten (Kapitel 4.2.1 und
4.2.2). In Kooperation mit der Arbeitsgruppe von R. Goforth (University of Arkansas)
wurden diese Daten in einer gemeinsamen Publikation zusammengestellt (Marty et al.,
2009, eingereicht). Ergebnisse aus der Arbeitsgruppe Goforth zeigten eine Copräzipitation
von rekombinantem cpSRP43 und Alb3. Diese Versuche erfolgten mit
Thylakoidmembranen aus Pisum sativum (Erbse), so dass es sich bei dem
copräzipitierten Alb3 um das Homolog PPF1 aus der Erbse handelte. Die Ansätze mit
rekombinantem cpSRP54 und cpFtsY zeigten sowohl einzeln, als auch in Kombination
kein copräzipitiertes Alb3 (PPF1). In den Ansätzen mit einer Kombination der
rekombinanten Proteine konnte nur in den Proben das Alb3 (PPF1) copräzipitiert werden,
in denen auch das cpSRP43 vorlag. Das cpSRP43 könnte somit als Brücke innerhalb der
cpSRP-Komponenten im Verlauf des docking-Prozesses an das Alb3 wirken.
Im Laufe dieser vorliegenden Doktorarbeit sind weitere Daten publiziert worden, die eine
Komplementation einer cpSRP43-defizienten chaos-Mutante von Arabidopsis thaliana mit
einem cpSRP43(ΔCD2) Protein zeigten (Tzvetkova-Chevolleau et al., 2007). Das
cpSRP43(ΔCD2) ist nicht mehr in der Lage mit dem cpSRP54 zu interagieren und einen
Transitkomplex mit dem cpSRP54 und LHCP zu bilden (Goforth et al., 2004). Aufgrund
der Komplementation wurde vermutet, dass cpSRP43 unabhängig von cpSRP54 den
LHCP-Transport vermitteln kann. Weiterhin wurde durch Immunlokalisationsstudien eine
Colokalisation von cpSRP43 mit der Translokase Alb3 gezeigt, die allerdings nur in
Abwesenheit von cpSRP54 zu beobachten war (Tzvetkova-Chevolleau et al., 2007). Die
direkte Interaktion der beiden Proteine wurde durch eine Copräzipitation des Alb3 mit
rekombinantem cpSRP43 gezeigt (Tzvetkova-Chevolleau et al., 2007). Die Autoren
vermuten daher einen alternativen Transportweg der LHCPs über cpSRP43 und Alb3, der
unabhängig von cpSRP54 und cpFtsY verläuft.
Pull down Analysen aus der vorliegenden Doktorarbeit zeigten allerdings, dass cpSRP43
sowohl allein, als auch im Komplex mit cpSRP54, mit Alb3 interagierte. Dieses Ergebnis
könnte einerseits die Möglichkeit eines alternativen Transportwegs unterstützen, bei dem
das cpSRP43 auch unabhängig von cpSRP54 den LHCP-Transport zur Translokase Alb3
vermittelt (Tzvetkova-Chevolleau et al., 2007). Andererseits ist aber bemerkenswert, dass
cpSRP43 auch im Komplex mit cpSRP54 und nicht nur in Abwesenheit des cpSRP54 mit
Alb3 interagierte.
5 Diskussion 105
5.2 Identifizierung der Alb3-Interaktionsbereiche für das cpSRP43 und Charakterisierung der Alb3-cpSRP43 Interaktion
Weiterführende Experimente sollten die Interaktionsdomäne des Alb3 für das cpSRP43
ermitteln. Als erster Ansatzpunkt sollten durch die Inkubation von salzgewaschenen und
proteaseverdauten Thylakoidmembranen mit den Proteinen cpSRP43, cpSRP54 und
cpFtsY mögliche stromale Bereiche des Alb3 als Interaktionsdomäne identifiziert werden
(Marty et al., 2009, eingereicht). Es zeigte sich, dass rekombinantes cpSRP43 und cpFtsY
an salzgewaschene Thylakoidmembranen von Arabidopsis thaliana binden konnten,
während eine Bindung des Kontrollproteins GST ausblieb. Ein Proteaseverdau der
Membranen führte zu einem vollständigen Verlust der Bindung des cpSRP43. Das cpFtsY
konnte jedoch, aufgrund seiner Membranaffinität, an die mit Protease behandelten
Thylakoidmembranen binden (Marty et al., 2009). Diese Ergebnisse wurden durch die
Bindungsversuche von radioaktiv markiertem cpSRP43, cpSRP54 und cpFtsY an
Thylakoidmembranen aus der Erbse bestätigt. Der Proteaseverdau der Membranen
bewirkte eine Abspaltung des stromalen C-Terminus von Alb3 (PPF1) (Marty et al., 2009,
eingereicht). Diese Ergebnisse deuteten darauf hin, dass die Bindung des cpSRP43 an
die Thylakoidmembranen über den stromalen C-terminus des Alb3 erfolgen könnte.
Weitere Daten aus der vorliegenden Doktorarbeit und der Kooperation (Marty et al., 2009,
eingereicht) zeigten, dass der C-Terminus von Alb3 tatsächlich direkt mit dem cpSRP43
interagierte, während keine Interaktion zwischen cpSRP54 und cpFtsY mit Alb3
nachgewiesen wurde. Ausführliche pull down Versuche mit rekombinantem cpSRP43 und
rekombinantem Alb3 C-Terminus (PPF1) bestätigten die Bedeutung des C-Terminus von
Alb3 für die Interaktion mit cpSRP43 (Marty et al., 2009, eingereicht). Die Messung der
Interaktion von cpSRP43 und dem C-Terminus von Alb3 mittels isothermaler
Titrationskalorimetrie bekräftigten, dass das cpSRP43 direkt mit dem löslichen C-
Terminus von Alb3 interagierte (Marty et al., 2009, eingereicht).
Ein Vergleich innerhalb der Oxa1/YidC/Alb3-Proteinfamilie zeigt, dass der C-Terminus für
die anderen homologen Proteine ebenfalls eine entscheidende Bedeutung als funktionale
Domäne hat. Das bakterielle Homolog YidC und das mitochondriale Homolog Oxa1 sind
in der Lage mit ihrem C-Terminus Ribosomen zu binden und damit die cotranslationale
Insertion von Membranproteinen zu ermöglichen (Szyrach et al., 2003; Kohler et al.,
2009). Es wird vermutet, dass das cpSRP43 im posttranslationalen cpSRP-Transportweg
anstelle von Ribosomen an das Alb3 bindet. Weitere Arbeiten zeigten, dass Alb3 mit der
cpSec-Translokase assoziiert ist und bei der cotranslationalen Insertion des plastidär
kodierten D1 beteiligt ist (Klostermann et al., 2002; Ossenbühl et al., 2004). Demnach
bleibt zu klären, in welcher Form und durch welchen Mechanismus sich der co- und
posttranslationale Import der Proteine über Alb3 in die Thylakoidmembran unterscheidet
5 Diskussion 106
und reguliert wird.
Der Nachweis der Interaktion von cpSRP43 und Alb3 stellte die Frage nach der
funktionellen Bedeutung und physiologischen Relevanz dieser Interaktion innerhalb des
posttranslationalen cpSRP-Transportwegs. Bisherige Arbeiten wiesen einen Einfluss der
GTP-Bindung und Hydrolyse der beiden GTPasen cpSRP54 und cpFtsY bei der
Integration der LHCPs nach (Tu et al., 1999; Moore et al., 2003). Zudem wurde ein
regulierender Effekt des cpSRP43 auf die GTP-Hydrolyse der GTPasen gezeigt (Goforth
et al., 2004). In Marty et al., 2009 (eingereicht) wird weiterhin gezeigt, dass das cpSRP43
die GTPase Aktivität des cpSRP54 und cpFtsY in Abhängigkeit von dem Alb3 C-Terminus
(PPF1) stimulierte. Daher wurde vermutet, dass die GTP-Hydrolyse innerhalb des cpSRP-
Transportwegs nur bei Anwesenheit der Translokase Alb3 erfolgt und dass die GTP-
Hydrolyse ein entscheidender Schritt für das Recycling der cpSRP-Komponenten ist. Das
cpSRP43 könnte damit als Vermittler zwischen dem Alb3, cpSRP54, cpFtsY und LHCP
dienen und die Interaktion zwischen cpSRP43 und Alb3 könnte eine regulatorische
Funktion an der Thylakoidmembran einnehmen.
Die Ergebnisse der vorliegenden Doktorarbeit deuten darauf hin, dass neben dem C-
Terminus noch die 5. Transmembrandomäne des Alb3 für eine Interaktion mit dem
cpSRP43 wichtig ist. Dieses wurde zunächst durch die bimolekulare
Fluoreszenzkomplementation gezeigt (Kapitel 4.2.3). Die Existenz einer neben dem C-
Terminus zusätzlichen Bindestelle für das cpSRP43 wurde durch das Peptide Scanning
unterstützt (Kapitel 4.2.4). So umfasste ein Cluster der detektierten PepSpots die
Aminosäuren 305-328 der 5. Transmembrandomäne von Alb3. Diese Spots zeigten eine
gemeinsame Sequenz der Aminosäuren 314-319 (LVFKFL). Die Identifizierung dieser
Aminosäuren war eine Bestätigung der Messung der Alb3-Alb4 overlap-Konstrukte im
Split-YFP System, bei der das Bindemotiv LVFKF (Aminosäuren 314-318) innerhalb des
Alb3 eine wesentliche Bedeutung für die Interaktion mit cpSRP43 hatte. Dieses Motiv
wurde weiterhin durch die Deletionen und Mutationen in diesem Sequenzmotiv des Alb3
als Bindestelle für das cpSRP43 im Split-Ubiquitin System und Split-YFP System bestätigt
(Kapitel 4.2.5). Erstaunlicherweise liegt diese Bindestelle auf der lumenalen Seite der
5. Transmembrandomäne von Alb3. Daher stellt sich die Frage nach der Zugänglichkeit
dieser Bindestelle für das stromale cpSRP43 innerhalb des cpSRP-Transportwegs.
Kürzlich veröffentlichte Daten haben gezeigt, dass die homologen Proteine YidC und
Oxa1 in Anwesenheit von translatierenden Ribosomen möglicherweise eine Pore bilden
(Kohler et al., 2009). Die Interaktion des YidC und Oxa1 mit den Ribosomen erfolgt über
den jeweiligen C-Terminus. Mit Hilfe von BN-PAGE und Crosslinking Studien wurde
gezeigt, dass die Ribosomen die dimere Form von YidC und Oxa1 stabilisieren. Weitere
Analysen ergaben, dass die Dimere von YidC und Oxa1 über dem Ausgang des
Ribosomentunnels lokalisiert vorliegen.
5 Diskussion 107
Mit der Existenz des Bindemotivs innerhalb der 5. Transmembrandomäne des Alb3
könnte ein Mechanismus vermutet werden, bei dem das Alb3 für den docking-Prozess
des Transitkomplexes und die Insertion der LHCPs in die Thylakoidmembran eine Pore
ausbildet. Die Topologie des Alb3 beruht jedoch bisher nur auf Strukturvorhersagen.
Daher kann nicht ausgeschlossen werden, dass die Bindestelle möglicherweise bei einer
abweichenden Struktur des Alb3 innerhalb der Thylakoidmembran für die Interaktion mit
dem cpSRP43 auch ohne Ausbildung einer Pore zugänglich wäre.
Die Ergebnisse der Messungen im Split-YFP ergaben außerdem, dass der C-Terminus
von Alb3, wie bereits beschrieben, für eine Interaktion mit dem cpSRP43 essentiell ist
(Kapitel 4.2.3). Über die Methode des Peptide Scanning wurden die Aminosäuren 374-
388 des Alb3 C-Terminus als Bindestelle für das cpSRP43 identifiziert (Kapitel 4.2.4). Die
Deletion dieser Bindestelle innerhalb des Alb3 führte sowohl im Split-YFP System, als
auch im Split-Ubiquitin System zum Verlust der Interaktion mit cpSRP43 (Kapitel 4.2.5
und 4.2.6). Die Bindestelle innerhalb des C-Terminus von Alb3 wurde durch den Einsatz
eines Alb3-Peptids (Aminosäuren 367-388), welches die Aminosäuren der potentiellen
Bindestelle (374-388) umfasste, bestätigt, da die Vorinkubation des cpSRP43 mit dem
Alb3-Peptid zu einer verringerten Bindung des cpSRP43 an die Thylakoidmembranen
führte (Kapitel 4.2.8).
Um zu untersuchen, ob die beiden Bindestellen des Alb3 für das cpSRP43 konserviert
vorliegen, wurde ein Alignment der Alb3 Homologe einiger Streptophyten in diesen
Sequenzbereichen durchgeführt (Abbildung 5-1). Phylogenetische Analysen (Zhang et al.,
2009) zeigten, dass während der Entwicklung der Landpflanzen von den Grünalgen eine
Kopie des Alb3 Gens verloren gegangen ist und eine Duplikation der anderen Kopie zu
zwei Genen geführt hat, welche die beiden Proteine Alb3 und Alb4 kodieren. In Populus
trichocarpa wurden diese beiden Gene erneut dupliziert, so dass es dort
dementsprechend vier Gene, zwei für Alb3 und zwei für Alb4, gibt.
Die identifizierte Bindestelle innerhalb der 5. Transmembrandomäne des Alb3 aus
Arabidopsis thaliana umfasst die Aminosäuren 314-318 (LVFKF). In dem
Sequenzvergleich der Alb3 Homologe dieser Streptophyten fällt auf, dass das Leucin (L)
an Position 314 sehr konserviert vorliegt (Abbildung 5-1). In dieser Arbeit wurde gezeigt,
dass Mutationen und Deletionen dieser Aminosäure des Alb3 in einem Verlust der
Bindung des cpSRP43 resultierten, während z. B. die Mutation des Phenylalanin an
Position 316 (F), welches innerhalb der Alb3 Proteine dieser Streptophyten nicht
konserviert vorliegt, keine Auswirkungen hatte. Die Aminosäuren Lysin und Phenylalanin
an Position 317 und 318 (KF) liegen ebenfalls sehr konserviert vor. Da diese
Aminosäuren allerdings auch in der Alb4 Sequenz vorkommen (Abbildung 5-2) und Alb4
keine Interaktion mit cpSRP43 zeigte, scheinen diese Aminosäuren innerhalb des LVFKF-
5 Diskussion 108
Motivs keine essentielle Bedeutung für die Bindung des cpSRP43 zu haben. Demzufolge
scheint in dem ersten Bindemotiv die Aminosäure Leucin an Position 314 für die Bindung
Abbildung 5-1: Alignment der C-terminalen Proteinsequenzen der Alb3 Homologe Die Abbildung zeigt ein Alignment der Proteinsequenzen der Alb3 Homologe im Bereich der 5.
Transmembrandomäne und des C-Terminus von Alb3 aus Arabidopsis thaliana. Die konservierten
Aminosäuren der Bindemotive im Bereich der Aminosäuren 314-318 und 374-388 sind in rot
dargestellt. Der Bereich der 5. Transmembrandomäne des Alb3 aus Arabidopsis thaliana ist grau
hinterlegt.
Die zweite Bindestelle im C-Terminus umfasst die Aminosäuren 374-388. Innerhalb dieser
identifizierten Bindestelle konnte ein konserviertes Motiv der Aminosäuren 375-377 (AKR)
gefolgt von einem Prolin an Position 382 innerhalb der gezeigten Alb3 Homologe der
Streptophyten erkannt werden (Abbildung 5-1). Interessanterweise ist dieser Bereich der
Bindestelle (Aminosäuren 374-388) und der etwas verlängerte Bereich des eingesetzten
synthetischen Peptids (Aminosäuren 367-388), welches zu einer Verringerung der
Bindung des cpSRP43 an die Thylakoidmembran führte, im Vergleich zu dem restlichen
5 Diskussion 109
C-Terminus dieser Streptophyten sehr homolog und konserviert. Diese Tatsache
bekräftigt somit die Bindestelle innerhalb des C-Terminus, da funktionelle Domänen
allgemein sehr konserviert vorliegen.
Das Alignment der Alb3 Sequenz aus Arabidopsis thaliana mit homologen Alb4 Proteinen
der Streptophyten zeigt, dass beide Motive der identifizierten Bindestellen für das
cpSRP43 in den Alb4 Proteinen nicht konserviert vorliegen (Abbildung 5-2). Dieses
unterstützt weiterhin das Ergebnis dieser Doktorarbeit, dass Alb4 nicht mit cpSRP43
interagierte. Dieses ist möglicherweise auf das Fehlen dieser in den Alb3 Proteinen
zeigten, dass die Heterodimerisierung des cpSRP-Komplexes auf einer Interaktion
zwischen der α-Helix in Chromodomäne 2 des cpSRP43 und einem 10 Aminosäuren
langen Bindemotiv innerhalb der M-Domäne des cpSRP54 beruht (Goforth et al., 2004;
Funke et al., 2005; Hermkes et al., 2006; Kathir et al., 2008). Die Bildung des
5 Diskussion 113
Transitkomplexes erfolgt durch Interaktion der Ankyrin repeat Domäne des cpSRP43 mit
einem konservierten sogenannten L18-Motiv des LHCPs (DeLille et al., 2000; Tu et al.,
2000; Jonas-Straube et al., 2001; Stengel et al., 2008). In dieser Doktorarbeit wurde die
Ankyrin repeat Region 4 des cpSRP43 als Interaktionsdomäne für die Translokase Alb3
identifiziert. Erstaunlicherweise stellt das cpSRP43 mit der Ankyrin repeat Region die
gleiche Binderegion für das Alb3, wie für das LHCP. In Stengel et al, 2008 wurde gezeigt,
dass das L18-Peptid des LHCP an die Oberfläche der Ankyrin repeats 2-4 bindet.
Weiterhin bewirkte eine Mutation des Tyrosin 204 der Ankyrin repeat Region 3 des
cpSRP43 einen Verlust der Bindung des L18-Peptids.
In diesem Zusammenhang stellt sich die Frage nach dem Zeitpunkt der Interaktion von
cpSRP43 und Alb3. Möglicherweise erfolgt die Interaktion des cpSRP43 und Alb3 erst zu
einem späteren Zeitpunkt der LHCP-Insertion. Das LHCP muss von den cpSRP-
Komponenten abgelöst und in die Membran integriert werden, bevor es zur Freisetzung
der Komponenten für nachfolgende Transportprozesse kommt. Daher ist es nötig den
Mechanismus der Freisetzung des LHCPs aus dem Transitkomplex für die Insertion in die
Thylakoidmembran zu analysieren. In Marty et al., 2009 (eingereicht) wurde gezeigt, dass
der Alb3 C-Terminus einen Effekt auf die Transitkomplexbildung zwischen cpSRP43,
cpSRP54 und LHCP hatte. In Anwesenheit des Alb3 C-Terminus kam es nicht zum
Einwandern des Transitkomplexes in ein nichtdenaturierendes Gel. Es konnte bereits
gezeigt werden, dass das LHCP nur in Form des Transitkomplexes in die Gelmatrix
einwanderte (DeLille et al., 2000). Das bedeutet, dass Alb3 einen spezifischen Effekt auf
die Stabilität des Transitkomplexes oder auf das Einwandern des Komplexes in das
nichtdenaturierende Gel hatte. Man nimmt an, dass Alb3 mit dem cpSRP43 des
Transitkomplexes interagieren kann, und dass diese Interaktion eine Bedeutung für die
Freisetzung und Insertion der LHCPs haben könnte. Messungen über die isothermale
Titrationskalorimetrie zeigten, dass nicht nur das cpSRP43, sondern auch der Alb3 C-
Terminus, das L18-Motiv des LHCPs bindet (Marty et al., 2009, eingereicht). Weitere
Analysen ergaben, dass zwischen dem Alb3 C-Terminus und cpSRP43 eine ähnliche
Bindungsaffinität vorlag wie zwischen dem Alb3 C-Terminus und einem vorgeformten
Komplex aus cpSRP43 und dem L18-Peptid (Marty et al., 2009, eingereicht).
Interessanterweise hatte ein vorgeformter Komplex aus dem Alb3 C-Terminus und
cpSRP43 eine geringere Bindungsaffinität für das L18-Peptid, als der Alb3 C-Terminus
und das cpSRP43 allein. Diese Beobachtung könnte so interpretiert werden, dass die
Interaktion zwischen Alb3 und cpSRP43 eine nachträgliche Bindung des LHCPs
verhindert, sobald es aus dem Transitkomplex freigesetzt wurde.
Die Bedeutung der Interaktion von Alb3 und cpSRP43 wird möglicherweise in dem
docking-Prozess des Transitkomplexes an die Thylakoidmembran, in der Regulation der
GTPase Aktivität des cpSRP54 und cpFtsY und in der Freisetzung der LHCPs für die
5 Diskussion 114
Insertion in die Membran liegen.
In Abbildung 5-4 ist ein Modell des posttranslationalen cpSRP-Transportwegs dargestellt.
Das LHCP wird von dem cpSRP43 und cpSRP54 gebunden und über den Transitkomplex
zum Thylakoidmembran assoziierten cpSRP-Rezeptor cpFtsY transportiert. An der
Thylakoidmembran kommt es zur Interaktion von Alb3 mit dem cpSRP43. Aufgrund dieser
Interaktion des membrangebundenen Komplexes mit der Translokase Alb3 erfolgt nach
GTP-Hydrolyse die Insertion der LHCPs und Freisetzung der löslichen cpSRP-
Komponenten.
Transit-komplex
LHCP
Thylakoidmembran
Alb3
cpSRP
cpFtsY43 54 43 54
43 54
43 54
43 54
TT
TT D
D
Abbildung 5-4: Modell des posttranslationalen cpSRP-Transportwegs Das cpSRP (chloroplast signal recognition particle) besteht aus den beiden Proteinkomponenten
cpSRP43 und cpSRP54 und bildet zusammen mit dem LHCP (light-harvesting chlorophyll-a/b-
binding protein) den Transitkomplex. Der Transitkomplex ermöglicht den Transport der
hydrophoben LHCPs durch das Stroma zur Thylakoidmembran und bildet mit dem cpSRP-
Rezeptor cpFtsY einen membrangebundenen Komplex. Nach erfolgter Interaktion von Alb3 mit
cpSRP43 kommt es nach GTP-Hydrolyse zur Insertion der LHCPs und zur Freisetzung der
löslichen cpSRP-Komponenten.
5 Diskussion 115
5.4 Alb4 interagiert nicht mit den Komponenten des cpSRP-Transportwegs
Das Alb4 wurde als zweites chloroplastidäres Homolog der Oxa1/YidC/Alb3-Proteinfamilie
in Arabidopsis thaliana identifiziert (Gerdes et al., 2006). GFP-Lokalisationsstudien und
Importstudien zeigten, dass Alb4 im Chloroplasten lokalisiert ist und als Membranprotein
vorliegt. Die Analyse chloroplastidärer Subfraktionen (Stroma, Hüllmembranen und
Thylakoidmembranen) ergab eine ausschließliche Lokalisation des Alb4 in der
Thylakoidmembran von Arabidopsis thaliana. Eine T-DNA Insertionsmutante und eine
RNAi-Mutante von Arabidopsis thaliana mit einem Alb4 Proteinlevel von weniger als 10 %
zeigten Defekte in der chloroplastidären Ultrastruktur, währenddessen diese Pflanzen
and peptide inhibition studies identified two binding sites of Alb3 for the cpSRP43. The
binding motif within the fifth transmembrane domain of Alb3 consists of the amino acids
314-318 (LVFKF) whilst the binding domain in the C-Terminus comprises the amino acids
374-388. Furthermore the binding domain of cpSRP43 for the interaction with Alb3 was
narrowed down. Thylakoid membrane binding studies suggested an essential role of
ankyrin repeat 4 for thylakoid membrane binding of cpSRP43.
The aim of the second part of the thesis was to provide an indication of the function of the
thylakoid membrane protein Alb4 by protein interaction studies. Crosslinking studies,
bimolecular fluorescence complementation studies and gelfiltration analyses revealed a
direct interaction and association of Alb4 with the chloroplast ATP-synthase. In addition
gelfiltration analyses and coimmunoprecipitation studies indicated that Alb4 is not
associated in the same complex as Alb3 and cpSecY. According to this contention, Alb4
seems to have an independent function of Alb3 and the cpSec-translocase in the insertion
and assembly of the ATP-synthase.
8 Literaturverzeichnis 122
8. Literaturverzeichnis
Abdallah F, Salamini F, Leister D (2000) A prediction of the size and evolutionary origin of the proteome of chloroplasts of Arabidopsis. Trends Plant Sci 5: 141-142
Agne B, Kessler F (2009) Protein transport in organelles: The Toc complex way of preprotein import. FEBS J 276: 1156-1165
Aldridge C, Cain P, Robinson C (2009) Protein transport in organelles: Protein transport into and across the thylakoid membrane. FEBS J 276: 1177-1186
Altamura N, Capitanio N, Bonnefoy N, Papa S, Dujardin G (1996) The Saccharomyces cerevisiae OXA1 gene is required for the correct assembly of cytochrome c oxidase and oligomycin-sensitive ATP synthase. FEBS Lett 382: 111-115
Bauer M, Behrens M, Esser K, Michaelis G, Pratje E (1994) PET1402, a nuclear gene required for proteolytic processing of cytochrome oxidase subunit 2 in yeast. Mol Gen Genet 245: 272-278
Beck K, Eisner G, Trescher D, Dalbey RE, Brunner J, Muller M (2001) YidC, an assembly site for polytopic Escherichia coli membrane proteins located in immediate proximity to the SecYE translocon and lipids. EMBO Rep 2: 709-714
Bellafiore S, Ferris P, Naver H, Göhre V, Rochaix JD (2002) Loss of Albino3 leads to the specific depletion of the light-harvesting system. Plant Cell 14: 2303-2314
Benz JP, Soll J, Bolter B (2009) Protein transport in organelles: The composition, function and regulation of the Tic complex in chloroplast protein import. FEBS J 276: 1166-1176
Bonnefoy N, Chalvet F, Hamel P, Slonimski PP, Dujardin G (1994) OXA1, a Saccharomyces cerevisiae nuclear gene whose sequence is conserved from prokaryotes to eukaryotes controls cytochrome oxidase biogenesis. J Mol Biol 239: 201-212
Bonnefoy N, Fiumera HL, Dujardin G, Fox TD (2009) Roles of Oxa1-related inner-membrane translocases in assembly of respiratory chain complexes. Biochim Biophys Acta 1793: 60-70
Braun NA, Davis AW, Theg SM (2007) The chloroplast Tat pathway utilizes the transmembrane electric potential as an energy source. Biophys J 93: 1993-1998
Brehm A, Tufteland KR, Aasland R, Becker PB (2004) The many colours of chromodomains. Bioessays 26: 133-140
Bruce BD (2000) Chloroplast transit peptides: structure, function and evolution. Trends Cell Biol 10: 440-447
Bruce BD (2001) The paradox of plastid transit peptides: conservation of function despite divergence in primary structure. Biochim Biophys Acta 1541: 2-21
Brundage L, Hendrick JP, Schiebel E, Driessen AJ, Wickner W (1990) The purified E. coli integral membrane protein SecY/E is sufficient for reconstitution of SecA-dependent precursor protein translocation. Cell 62: 649-657
Bullock WO, Fernandez JM, Short JM (1987). XL1-Blue: a high efficiency plasmid transforming recA Escherichia coli strain with beta-glucosidase selection. Biotechnique 5: 376 378.
Cavalier-Smith T (2000) Membrane heredity and early chloroplast evolution. Trends Plant Sci 5: 174-182
8 Literaturverzeichnis 123
Chaddock AM, Mant A, Karnauchov I, Brink S, Herrmann RG, Klösgen RB, Robinson C (1995) A new type of signal peptide: central role of a twin-arginine motif in transfer signals for the delta pH-dependent thylakoidal protein translocase. EMBO J 14: 2715-2722
Chandrasekar S, Chartron J, Jaru-Ampornpan P, Shan SO (2008) Structure of the chloroplast signal recognition particle (SRP) receptor: domain arrangement modulates SRP-receptor interaction. J Mol Biol 375: 425-436
Chen M, Samuelson JC, Jiang F, Muller M, Kuhn A, Dalbey RE (2002) Direct interaction of YidC with the Sec-independent Pf3 coat protein during its membrane protein insertion. J Biol Chem 277: 7670-7675
Cline K, Mori H (2001) Thylakoid DeltapH-dependent precursor proteins bind to a cpTatC-Hcf106 complex before Tha4-dependent transport. J Cell Biol 154: 719-729
Dabney-Smith C, Mori H, Cline K (2006) Oligomers of Tha4 organize at the thylakoid Tat translocase during protein transport. J Biol Chem 281: 5476-5483
de Gier JW, Luirink J (2001) Biogenesis of inner membrane proteins in Escherichia coli. Mol Microbiol 40: 314-322
DeLille J, Peterson EC, Johnson T, Moore M, Kight A, Henry R (2000) A novel precursor recognition element facilitates posttranslational binding to the signal recognition particle in chloroplasts. Proc Natl Acad Sci U S A 97: 1926-1931
Di Cola A, Bailey S, Robinson C (2005) The thylakoid delta pH/delta psi are not required for the initial stages of Tat-dependent protein transport in tobacco protoplasts. J Biol Chem 280: 41165-41170
Driessen AJ, Nouwen N (2008) Protein translocation across the bacterial cytoplasmic membrane. Annu Rev Biochem 77: 643-667
Facey SJ, Neugebauer SA, Krauss S, Kuhn A (2007) The mechanosensitive channel protein MscL is targeted by the SRP to the novel YidC membrane insertion pathway of Escherichia coli. J Mol Biol 365: 995-1004
Finazzi G, Chasen C, Wollman FA, de Vitry C (2003) Thylakoid targeting of Tat passenger proteins shows no delta pH dependence in vivo. EMBO J 22: 807-815
Franklin AE, Hoffman NE (1993). Characterization of a Chloroplast Homologue of the 54-kDa Subunit of the Signal Recognition Particle. J Biol Chem 268: 22175-22180.
Funes S, Hasona A, Bauerschmitt H, Grubbauer C, Kauff F, Collins R, Crowley PJ, Palmer SR, Brady LJ, Herrmann JM (2009) Independent gene duplications of the YidC/Oxa/Alb3 family enabled a specialized cotranslational function. Proc Natl Acad Sci U S A 106: 6656-6661
Funes S, Nargang FE, Neupert W, Herrmann JM (2004) The Oxa2 protein of Neurospora crassa plays a critical role in the biogenesis of cytochrome oxidase and defines a ubiquitous subbranch of the Oxa1/YidC/Alb3 protein family. Mol Biol Cell 15: 1853-1861
Funke S, Knechten T, Ollesch J, Schünemann D (2005) A unique sequence motif in the 54-kDa subunit of the chloroplast signal recognition particle mediates binding to the 43-kDa subunit. J Biol Chem 280: 8912-8917
Gathmann S, Eva R, Uwe K, Schneider D (2008) A conserved structure and function of the YidC homologous protein Slr1471 from Synechocystis sp. PCC 6803. J Microbiol Biotechnol 18: 1090-1094
8 Literaturverzeichnis 124
Gerdes L, Bals T, Klostermann E, Karl M, Philippar K, Hunken M, Soll J, Schünemann D (2006) A second thylakoid membrane-localized Alb3/OxaI/YidC homologue is involved in proper chloroplast biogenesis in Arabidopsis thaliana. J Biol Chem 281: 16632-16642
Glick BS, Von Heijne G (1996) Saccharomyces cerevisiae mitochondria lack a bacterial-type sec machinery. Protein Sci 5: 2651-2652
Goforth RL, Peterson EC, Yuan J, Moore MJ, Kight AD, Lohse MB, Sakon J, Henry RL (2004) Regulation of the GTPase cycle in post-translational signal recognition particle-based protein targeting involves cpSRP43. J Biol Chem 279: 43077-43084
Göhre V, Ossenbühl F, Crevecoeur M, Eichacker LA, Rochaix JD (2006) One of two alb3 proteins is essential for the assembly of the photosystems and for cell survival in Chlamydomonas. Plant Cell 18: 1454-1466
Groves MR, Mant A, Kuhn A, Koch J, Dubel S, Robinson C, Sinning I (2001) Functional characterization of recombinant chloroplast signal recognition particle. J Biol Chem 276: 27778-27786
Hell K, Herrmann JM, Pratje E, Neupert W, Stuart RA (1998) Oxa1p, an essential component of the N-tail protein export machinery in mitochondria. Proc Natl Acad Sci U S A 95: 2250-2255
Hell K, Neupert W, Stuart RA (2001) Oxa1p acts as a general membrane insertion machinery for proteins encoded by mitochondrial DNA. EMBO J 20: 1281-1288
Hermkes R, Funke S, Richter C, Kuhlmann J, Schünemann D (2006) The alpha-helix of the second chromodomain of the 43 kDa subunit of the chloroplast signal recognition particle facilitates binding to the 54 kDa subunit. FEBS Lett 580: 3107-3111
Hoffman NE, Franklin AE (1994) Evidence for a stromal GTP requirement for the integration of a chlorophyll a/b-binding polypeptide into thylakoid membranes. Plant Physiol 105: 295-304
Houben EN, ten Hagen-Jongman CM, Brunner J, Oudega B, Luirink J (2004) The two membrane segments of leader peptidase partition one by one into the lipid bilayer via a Sec/YidC interface. EMBO Rep 5: 970-975
Jackson-Constan D, Akita M, Keegstra K (2001) Molecular chaperones involved in chloroplast protein import. Biochim Biophys Acta 1541: 102-113
Jaru-Ampornpan P, Chandrasekar S, Shan SO (2007) Efficient interaction between two GTPases allows the chloroplast SRP pathway to bypass the requirement for an SRP RNA. Mol Biol Cell 18: 2636-2645
Jarvis P (2008) Targeting of nucleus-encoded proteins to chloroplasts in plants. New Phytol 179: 257-285
Jarvis P, Robinson C (2004) Mechanisms of protein import and routing in chloroplasts. Curr Biol 14: R1064-1077
Jia L, Dienhart M, Schramp M, McCauley M, Hell K, Stuart RA (2003) Yeast Oxa1 interacts with mitochondrial ribosomes: the importance of the C-terminal region of Oxa1. EMBO J 22: 6438-6447
Jia L, Dienhart MK, Stuart RA (2007) Oxa1 directly interacts with Atp9 and mediates its assembly into the mitochondrial F1Fo-ATP synthase complex. Mol Biol Cell 18: 1897-1908
8 Literaturverzeichnis 125
Jiang F, Yi L, Moore M, Chen M, Rohl T, Van Wijk KJ, De Gier JW, Henry R, Dalbey RE (2002) Chloroplast YidC homolog Albino3 can functionally complement the bacterial YidC depletion strain and promote membrane insertion of both bacterial and chloroplast thylakoid proteins. J Biol Chem 277: 19281-19288
Jonas-Straube E, Hutin C, Hoffman NE, Schünemann D (2001) Functional analysis of the protein-interacting domains of chloroplast SRP43. J Biol Chem 276: 24654-24660
Kathir KM, Rajalingam D, Sivaraja V, Kight A, Goforth RL, Yu C, Henry R, Kumar TK (2008) Assembly of chloroplast signal recognition particle involves structural rearrangement in cpSRP43. J Mol Biol 381: 49-60
Keegstra K, Cline K (1999) Protein import and routing systems of chloroplasts. Plant Cell 11: 557-570
Kessler F, Schnell DJ (2006) The function and diversity of plastid protein import pathways: a multilane GTPase highway into plastids. Traffic 7: 248-257
Kiefer D, Kuhn A (2007) YidC as an essential and multifunctional component in membrane protein assembly. Int Rev Cytol 259: 113-138
Kim SJ, Jansson S, Hoffman NE, Robinson C, Mant A (1999) Distinct "assisted" and "spontaneous" mechanisms for the insertion of polytopic chlorophyll-binding proteins into the thylakoid membrane. J Biol Chem 274: 4715-4721
Klimyuk VI, Persello-Cartieaux F, Havaux M, Contard-David P, Schünemann D, Meiherhoff K, Gouet P, Jones JD, Hoffman NE, Nussaume L (1999) A chromodomain protein encoded by the arabidopsis CAO gene is a plant-specific component of the chloroplast signal recognition particle pathway that is involved in LHCP targeting. Plant Cell 11: 87-99
Klösgen RB, Brock IW, Herrmann RG, Robinson C (1992) Proton gradient-driven import of the 16 kDa oxygen-evolving complex protein as the full precursor protein by isolated thylakoids. Plant Mol Biol 18: 1031-1034
Klostermann E, Droste Gen Helling I, Carde JP, Schünemann D (2002) The thylakoid membrane protein ALB3 associates with the cpSecY-translocase in Arabidopsis thaliana. Biochem J 368: 777-781
Koch HG, Moser M, Muller M (2003) Signal recognition particle-dependent protein targeting, universal to all kingdoms of life. Rev Physiol Biochem Pharmacol 146: 55-94
Kogata N, Nishio K, Hirohashi T, Kikuchi S, Nakai M (1999) Involvement of a chloroplast homologue of the signal recognition particle receptor protein, FtsY, in protein targeting to thylakoids. FEBS Lett 447: 329-333
Kohler R, Boehringer D, Greber B, Bingel-Erlenmeyer R, Collinson I, Schaffitzel C, Ban N (2009) YidC and Oxa1 form dimeric insertion pores on the translating ribosome. Mol Cell 34: 344-353
Kuhn A, Stuart R, Henry R, Dalbey RE (2003) The Alb3/Oxa1/YidC protein family: membrane-localized chaperones facilitating membrane protein insertion? Trends Cell Biol 13: 510-516
Laidler V, Chaddock AM, Knott TG, Walker D, Robinson C (1995) A SecY homolog in Arabidopsis thaliana. Sequence of a full-length cDNA clone and import of the precursor protein into chloroplasts. J Biol Chem 270: 17664-17667
Leister D (2003) Chloroplast research in the genomic age. Trends Genet 19: 47-56
8 Literaturverzeichnis 126
Li X, Henry R, Yuan J, Cline K, Hoffman NE (1995) A chloroplast homologue of the signal recognition particle subunit SRP54 is involved in the posttranslational integration of a protein into thylakoid membranes. Proc Natl Acad Sci U S A 92: 3789-3793
Marques JP, Schattat MH, Hause G, Dudeck I, Klösgen RB (2004) In vivo transport of folded EGFP by the DeltapH/TAT-dependent pathway in chloroplasts of Arabidopsis thaliana. J Exp Bot 55: 1697-1706
Martin W, Herrmann RG (1998) Gene transfer from organelles to the nucleus: how much, what happens, and Why? Plant Physiol 118: 9-17
Marty NJ, Rajalingam D, Kight AD, Lewis NE, Fologea D, Kumar TK, Henry RL, Goforth RL (2009) The membrane-binding motif of the chloroplast signal recognition particle receptor (cpFtsY) regulates GTPase activity. J Biol Chem 284: 14891-14903
May T, Soll J (2000) 14-3-3 proteins form a guidance complex with chloroplast precursor proteins in plants. Plant Cell 12: 53-64
Michl D, Robinson C, Shackleton JB, Herrmann RG, Klösgen RB (1994) Targeting of proteins to the thylakoids by bipartite presequences: CFoII is imported by a novel, third pathway. EMBO J 13: 1310-1317
Moore M, Goforth RL, Mori H, Henry R (2003) Functional interaction of chloroplast SRP/FtsY with the ALB3 translocase in thylakoids: substrate not required. J Cell Biol 162: 1245-1254
Moore M, Harrison MS, Peterson EC, Henry R (2000) Chloroplast Oxa1p homolog albino3 is required for post-translational integration of the light harvesting chlorophyll-binding protein into thylakoid membranes. J Biol Chem 275: 1529-1532
Mori H, Cline K (2002) A twin arginine signal peptide and the pH gradient trigger reversible assembly of the thylakoid [Delta]pH/Tat translocase. J Cell Biol 157: 205-210
Mori H, Summer EJ, Ma X, Cline K (1999) Component specificity for the thylakoidal Sec and Delta pH-dependent protein transport pathways. J Cell Biol 146: 45-56
Nilsson R, Brunner J, Hoffman NE, van Wijk KJ (1999) Interactions of ribosome nascent chain complexes of the chloroplast-encoded D1 thylakoid membrane protein with cpSRP54. EMBO J 18: 733-742
Nilsson R, van Wijk KJ (2002) Transient interaction of cpSRP54 with elongating nascent chains of the chloroplast-encoded D1 protein; 'cpSRP54 caught in the act'. FEBS Lett 524: 127-133
Ossenbühl F, Göhre V, Meurer J, Krieger-Liszkay A, Rochaix JD, Eichacker LA (2004) Efficient assembly of photosystem II in Chlamydomonas reinhardtii requires Alb3.1p, a homolog of Arabidopsis ALBINO3. Plant Cell 16: 1790-1800
Ossenbühl F, Inaba-Sulpice M, Meurer J, Soll J, Eichacker LA (2006) The synechocystis sp PCC 6803 oxa1 homolog is essential for membrane integration of reaction center precursor protein pD1. Plant Cell 18: 2236-2246
Pasch JC, Nickelsen J, Schünemann D (2005) The yeast split-ubiquitin system to study chloroplast membrane protein interactions. Appl Microbiol Biotechnol 69: 440-447
Payan LA, Cline K (1991) A stromal protein factor maintains the solubility and insertion competence of an imported thylakoid membrane protein. J Cell Biol 112: 603-613
8 Literaturverzeichnis 127
Pool MR (2005) Signal recognition particles in chloroplasts, bacteria, yeast and mammals (review). Mol Membr Biol 22: 3-15
Preuss M, Ott M, Funes S, Luirink J, Herrmann JM (2005) Evolution of mitochondrial oxa proteins from bacterial YidC. Inherited and acquired functions of a conserved protein insertion machinery. J Biol Chem 280: 13004-13011
Ravaud S, Stjepanovic G, Wild K, Sinning I (2008) The crystal structure of the periplasmic domain of the Escherichia coli membrane protein insertase YidC contains a substrate binding cleft. J Biol Chem 283: 9350-9358
Reyes-Prieto A, Weber AP, Bhattacharya D (2007) The origin and establishment of the plastid in algae and plants. Annu Rev Genet 41: 147-168
Richter S, Lamppa GK (1998) A chloroplast processing enzyme functions as the general stromal processing peptidase. Proc Natl Acad Sci U S A 95: 7463-7468
Richter ML, Hein R, Huchzermeyer B (2000) Important subunit interactions in the chloroplast ATP synthase. Biochim Biophys Acta 1458: 326-342
Robson A, Collinson I (2006) The structure of the Sec complex and the problem of protein translocation. EMBO Rep 7: 1099-1103
Roy LM, Barkan A (1998) A SecY homologue is required for the elaboration of the chloroplast thylakoid membrane and for normal chloroplast gene expression. J Cell Biol 141: 385-395
Saaf A, Monne M, de Gier JW, von Heijne G (1998) Membrane topology of the 60-kDa Oxa1p homologue from Escherichia coli. J Biol Chem 273: 30415-30418
Samuelson JC, Chen M, Jiang F, Moller I, Wiedmann M, Kuhn A, Phillips GJ, Dalbey RE (2000) YidC mediates membrane protein insertion in bacteria. Nature 406: 637-641
Saracco SA, Fox TD (2002) Cox18p is required for export of the mitochondrially encoded Saccharomyces cerevisiae Cox2p C-tail and interacts with Pnt1p and Mss2p in the inner membrane. Mol Biol Cell 13: 1122-1131
Schünemann D, Amin P, Hartmann E, Hoffman NE (1999) Chloroplast SecY is complexed to SecE and involved in the translocation of the 33-kDa but not the 23-kDa subunit of the oxygen-evolving complex. J Biol Chem 274: 12177-12182
Schünemann D, Gupta S, Persello-Cartieaux F, Klimyuk VI, Jones JD, Nussaume L, Hoffman NE (1998) A novel signal recognition particle targets light-harvesting proteins to the thylakoid membranes. Proc Natl Acad Sci U S A 95: 10312-10316
Schünemann D (2004) Structure and function of the chloroplast signal recognition particle. Curr Genet 44: 295-304
Schünemann D (2007) Mechanisms of protein import into thylakoids of chloroplasts. Biol Chem 388: 907-915
Scotti PA, Urbanus ML, Brunner J, de Gier JW, von Heijne G, van der Does C, Driessen AJ, Oudega B, Luirink J (2000) YidC, the Escherichia coli homologue of mitochondrial Oxa1p, is a component of the Sec translocase. EMBO J 19: 542-549
Sedgwick SG, Smerdon SJ (1999) The ankyrin repeat: a diversity of interactions on a common structural framework. Trends Biochem Sci 24: 311-316
Serek J, Bauer-Manz G, Struhalla G, van den Berg L, Kiefer D, Dalbey R, Kuhn A (2004) Escherichia coli YidC is a membrane insertase for Sec-independent proteins. EMBO J 23: 294-301
8 Literaturverzeichnis 128
Sivaraja V, Kumar TK, Leena PS, Chang AN, Vidya C, Goforth RL, Rajalingam D, Arvind K, Ye JL, Chou J, Henry R, Yu C (2005) Three-dimensional solution structures of the chromodomains of cpSRP43. J Biol Chem 280: 41465-41471
Soll J, Schleiff E (2004) Protein import into chloroplasts. Nat Rev Mol Cell Biol 5: 198-208
Spence E, Bailey S, Nenninger A, Moller SG, Robinson C (2004) A homolog of Albino3/OxaI is essential for thylakoid biogenesis in the cyanobacterium Synechocystis sp. PCC6803. J Biol Chem 279: 55792-55800
Stengel KF, Holdermann I, Cain P, Robinson C, Wild K, Sinning I (2008) Structural basis for specific substrate recognition by the chloroplast signal recognition particle protein cpSRP43. Science 321: 253-256
Stengel KF, Holdermann I, Wild K, Sinning I (2007) The structure of the chloroplast signal recognition particle (SRP) receptor reveals mechanistic details of SRP GTPase activation and a conserved membrane targeting site. FEBS Lett 581: 5671-5676
Sun C, Rusch SL, Kim J, Kendall DA (2007) Chloroplast SecA and Escherichia coli SecA have distinct lipid and signal peptide preferences. J Bacteriol 189: 1171-1175
Sundberg E, Slagter JG, Fridborg I, Cleary SP, Robinson C, Coupland G (1997) ALBINO3, an Arabidopsis nuclear gene essential for chloroplast differentiation, encodes a chloroplast protein that shows homology to proteins present in bacterial membranes and yeast mitochondria. Plant Cell 9: 717-730
Szyrach G, Ott M, Bonnefoy N, Neupert W, Herrmann JM (2003) Ribosome binding to the Oxa1 complex facilitates co-translational protein insertion in mitochondria. EMBO J 22: 6448-6457
Theg SM, Cline K, Finazzi G, Wollman FA (2005) The energetics of the chloroplast Tat protein transport pathway revisited. Trends Plant Sci 10: 153-154
Thompson SJ, Kim SJ, Robinson C (1998) Sec-independent insertion of thylakoid membrane proteins. Analysis of insertion forces and identification of a loop intermediate involving the signal peptide. J Biol Chem 273: 18979-18983
Tu CJ, Peterson EC, Henry R, Hoffman NE (2000) The L18 domain of light-harvesting chlorophyll proteins binds to chloroplast signal recognition particle 43. J Biol Chem 275: 13187-13190
Tu CJ, Schünemann D, Hoffman NE (1999) Chloroplast FtsY, chloroplast signal recognition particle, and GTP are required to reconstitute the soluble phase of light-harvesting chlorophyll protein transport into thylakoid membranes. J Biol Chem 274: 27219-27224
Tzvetkova-Chevolleau T, Hutin C, Noel LD, Goforth R, Carde JP, Caffarri S, Sinning I, Groves M, Teulon JM, Hoffman NE, Henry R, Havaux M, Nussaume L (2007) Canonical signal recognition particle components can be bypassed for posttranslational protein targeting in chloroplasts. Plant Cell 19: 1635-1648
8 Literaturverzeichnis 129
Urbanus ML, Scotti PA, Froderberg L, Saaf A, de Gier JW, Brunner J, Samuelson JC, Dalbey RE, Oudega B, Luirink J (2001) Sec-dependent membrane protein insertion: sequential interaction of nascent FtsQ with SecY and YidC. EMBO Rep 2: 524-529
van Bloois E, Koningstein G, Bauerschmitt H, Herrmann JM, Luirink J (2007) Saccharomyces cerevisiae Cox18 complements the essential Sec-independent function of Escherichia coli YidC. FEBS J 274: 5704-5713
van Bloois E, Nagamori S, Koningstein G, Ullers RS, Preuss M, Oudega B, Harms N, Kaback HR, Herrmann JM, Luirink J (2005) The Sec-independent function of Escherichia coli YidC is evolutionary-conserved and essential. J Biol Chem 280: 12996-13003
van der Laan M, Bechtluft P, Kol S, Nouwen N, Driessen AJ (2004) F1F0 ATP synthase subunit c is a substrate of the novel YidC pathway for membrane protein biogenesis. J Cell Biol 165: 213-222
van der Laan M, Houben EN, Nouwen N, Luirink J, Driessen AJ (2001) Reconstitution of Sec-dependent membrane protein insertion: nascent FtsQ interacts with YidC in a SecYEG-dependent manner. EMBO Rep 2: 519-523
Vörös (2007) Proteinintegration in die Thylakoidmembran von Arabidopsis thaliana- Untersuchung von Protein-Protein-Interaktionen des integralen Membranproteins Albino3 (Masterarbeit, Ruhr-Universität Bochum)
Walter M, Chaban C, Schutze K, Batistic O, Weckermann K, Nake C, Blazevic D, Grefen C, Schumacher K, Oecking C, Harter K, Kudla J (2004) Visualization of protein interactions in living plant cells using bimolecular fluorescence complementation. Plant J 40: 428-438
Weeden NF (1981) Genetic and biochemical implications of the endosymbiotic origin of the chloroplast. J Mol Evol 17: 133-139
Woolhead CA, Thompson SJ, Moore M, Tissier C, Mant A, Rodger A, Henry R, Robinson C (2001) Distinct Albino3-dependent and -independent pathways for thylakoid membrane protein insertion. J Biol Chem 276: 40841-40846
Xie K, Dalbey RE (2008) Inserting proteins into the bacterial cytoplasmic membrane using the Sec and YidC translocases. Nat Rev Microbiol 6: 234-244
Yamamoto HY, Bassi R (1996). Carotenoids: Localization and function. In Oxygenic Photosynthesis: The Light Reactions, D.R. Ort and C.F. Yocum, eds (Dordrecht, The Netherlands: Kluwer Academic Publishers) 539-563
Yen MR, Harley KT, Tseng YH, Saier MH, Jr. (2001) Phylogenetic and structural analyses of the oxa1 family of protein translocases. FEMS Microbiol Lett 204: 223-231
Yi L, Dalbey RE (2005) Oxa1/Alb3/YidC system for insertion of membrane proteins in mitochondria, chloroplasts and bacteria (review). Mol Membr Biol 22: 101-111
Yuan J, Henry R, McCaffery M, Cline K (1994) SecA homolog in protein transport within chloroplasts: evidence for endosymbiont-derived sorting. Science 266: 796-798
Yuan J, Kight A, Goforth RL, Moore M, Peterson EC, Sakon J, Henry R (2002) ATP stimulates signal recognition particle (SRP)/FtsY-supported protein integration in chloroplasts. J Biol Chem 277: 32400-32404
8 Literaturverzeichnis 130
Zhang L, Paakkarinen V, Suorsa M, Aro EM (2001) A SecY homologue is involved in chloroplast-encoded D1 protein biogenesis. J Biol Chem 276: 37809-37814
Zhang YJ, Tian HF, Wen JF (2009) The evolution of YidC/Oxa/Alb3 family in the three domains of life: a phylogenomic analysis. BMC Evol Biol 9: 137
Zygadlo A, Robinson C, Scheller HV, Mant A, Jensen PE (2006) The properties of the positively charged loop region in PSI-G are essential for its "spontaneous" insertion into thylakoids and rapid assembly into the photosystem I complex. J Biol Chem 281: 10548-10554
9 Abkürzungsverzeichnis 131
9. Abkürzungsverzeichnis
Ala Alanin
Alb3 Albino3
Ank Ankyrin repeat
Aqua dest. lat. Aqua destillata
AS Aminosäure(n)
ATP Adenosintriphosphat
BCA Bicinchoninsäure
bp Basenpaar(e)
BSA engl. bovine serum albumin
CD Chromodomäne
cpSRP engl. chloroplast signal recognition particle
DEPC engl. Diethylpyrocarbonate
DMSO Dimethylsulfoxid
DNA Desoxyribonukleinsäure
E. coli Escherichia coli
EDTA Ethylendiamintetraacetat
GST Glutathion S-Transferase
GTP Guanosintriphosphat
His Histidin
IPTG Isopropyl-β-D-thiogalactopyranosid
kDa kilo-Dalton
LHCP engl. light harvesting chlorophyll a/b binding protein
mRNA messenger-RNA
Ni-NTA engl. Nickel-nitrilotriacetic acid
NTP Nukleosidtriphosphat
OD Optische Dichte
PAGE Polyacrylamid-Gelelektrophorese
RNA Ribonukleinsäure
rpm engl. rotations per minute
SDS engl. sodium dodecyl sulfate
TEMED N,N,N’,N’,-Tetramethylethylendiamin
UV ultraviolett
YFP engl. yellow fluorescent protein
Weitere allgemein gebräuchliche Abkürzungen und Maßeinheiten sind nicht gesondert
aufgeführt.
10 Abbildungsverzeichnis 132
10. Abbildungsverzeichnis
Abbildung 1-1: Transportwege der Proteine innerhalb des Chloroplasten......................3 Abbildung 1-2: Modell des co- und posttranslationalen cpSRP-Transportwegs .............7 Abbildung 1-3: Proteininteraktionen der Komponenten des cpSRP-Transportwegs ......9 Abbildung 1-4: Membrantopologie von Alb3, YidC und Oxa1.......................................11 Abbildung 1-5: Proteininsertionen durch Alb3, YidC und Oxa1 ....................................16 Abbildung 4-1: Alignment der Aminosäuresequenz von Alb3 und Alb4........................64 Abbildung 4-2: Test der Antikörper und Präimmunseren gegen die Antigene GST-
cAlb3 und cAlb4-his ............................................................................66 Abbildung 4-3: Test der Antikörper gegen die in vitro translatierten
Volllängenproteine ..............................................................................67 Abbildung 4-4: Test der Antikörper gegen Thylakoidmembranproteine........................68 Abbildung 4-5: Alb3 interagiert mit cpSRP43 in Arabidopsis thaliana Protoplasten .....73 Abbildung 4-6: Expressionsnachweis der YFP-Fusionsproteine cpSRP43-nYFP,
Alb3-cYFP und Alb4-cYFP..................................................................75 Abbildung 4-7: Schematische Darstellung der vorhergesagten Membrantopologie
von Alb3 und Alb4 ...............................................................................76 Abbildung 4-8: Identifizierung der Alb3 Bindestelle für das cpSRP43 in Arabidopsis
Protoplasten ........................................................................................78 Abbildung 4-9: Alignment der YFP-Fusionsproteine zur Identifizierung der
Bindestelle...........................................................................................79 Abbildung 4-10: Peptide Scanning von Alb3 Peptiden mit rekombinantem cpSRP43..81 Abbildung 4-11: Alignment von Alb3 und Alb4 im Bereich des Bindemotivs ................82 Abbildung 4-12: Expressionsnachweis ausgewählter Alb3 Deletions- und
Mutationskonstrukte in Arabidopsis Protoplasten ...............................84 Abbildung 4-13: Pull down Analysen von rekombinantem cpSRP43 und cpSRP54
mit in vitro Translationsprodukten von Alb3 und Alb4 .........................88 Abbildung 4-14: Einfluss eines synthetischen Peptids von Alb3 auf die
Thylakoidmembranbindung von cpSRP43..........................................90 Abbildung 4-15: Schematische Darstellung des cpSRP43 ...........................................91 Abbildung 4-16: Thylakoidmembran-Bindungstest mit GST-cpSRP43
Deletionsproteinen (I)..........................................................................92 Abbildung 4-17: Thylakoidmembran-Bindungstest mit GST-cpSRP43
Deletionsproteinen (II).........................................................................94 Abbildung 4-18: Crosslinking von Alb4 und CF1β .........................................................96 Abbildung 4-19: Alb4 interagiert mit der ATP-Synthase Untereinheit CF0II in
Arabidopsis thaliana Protoplasten.......................................................98 Abbildung 4-20: Expressionsnachweis der YFP-Fusionsproteine Alb4-cYFP und
CF0II-nYFP..........................................................................................99 Abbildung 4-21: Alb4 cofraktioniert mit den ATP-Synthase Untereinheiten................100 Abbildung 4-22: Coimmunpräzipitation von Alb3, Alb4 und cpSecY...........................101 Abbildung 5-1: Alignment der C-terminalen Proteinsequenzen der Alb3 Homologe ..108
10 Abbildungsverzeichnis 133
Abbildung 5-2: Alignment des Alb3 aus Arabidopsis thaliana mit den Alb4 Proteinen anderer Streptophyten ......................................................................109
Abbildung 5-3: Proteininteraktionen des cpSRP43.....................................................112 Abbildung 5-4: Modell des posttranslationalen cpSRP-Transportwegs ......................114
11 Tabellenverzeichnis 134
11. Tabellenverzeichnis
Tabelle 3-1: Chemikalienliste ........................................................................................18 Tabelle 3-2: Verbrauchsmaterial ...................................................................................19 Tabelle 3-3: Kits ............................................................................................................19 Tabelle 3-4: Geräte .......................................................................................................20 Tabelle 3-5: Verwendete Plasmide ...............................................................................22 Tabelle 3-6: Escherichia coli-Stämme...........................................................................29 Tabelle 3-7: Saccharomyces cerevisiae-Stamm...........................................................29 Tabelle 3-8: Trenngel pH 8,8 (4 Minigele):....................................................................46 Tabelle 4-1: Alb3 interagiert mit cpSRP43 im Split-Ubiquitin System ...........................71 Tabelle 4-2: Interaktionstests im Split-YFP System......................................................74 Tabelle 4-3: Negativkontrollen der Interaktionstests im Split-YFP System ...................74 Tabelle 4-4: Interaktion der Alb3 Deletions- und Mutationsproteine mit cpSRP43-
nYFP in Arabidopsis Protoplasten ............................................................83 Tabelle 4-5: Interaktionstest von Alb3 Deletionsproteinen mit cpSRP43 im Hefe
Split-Ubiquitin System ...............................................................................85
12 Anhang 135
12. Anhang
12.1 Publikationen
Naomi J. Marty*, Nathaniel E. Lewis*, Thomas Bals*, Karuppanan Muthusamy Kathir, Dakshinamurthy Rajalingam, Alicia D. Kight, Beatrix Dünschede, Aniko Vörös, Thallapuranam Krishnaswamy Suresh Kumar, Ralph L. Henry, Danja Schünemann, and Robyn L. Goforth A Dynamic CpSRP43-Alb3 Interaction Mediates
Translocase Regulation of CpSRP Targeting Components; eingereicht * gleichberechtigte Erstautoren
Benz M., Bals T., Gügel I.L., Piotrowski M., Kuhn A., Schünemann D., Soll J., Ankele E. (2009) Alb4 of Arabidopsis promotes assembly and stabilisation of a none
chlorophyll-binding photosynthetic complex, the CF1CF0-ATP synthase. Mol. Plant; im
Druck
Gerdes L*, Bals T*, Klostermann E, Karl M, Philippar K, Hunken M, Soll J, Schünemann D (2006) A second thylakoid membrane-localized Alb3/OxaI/YidC
homologue is involved in proper chloroplast biogenesis in Arabidopsis thaliana. J Biol Chem 281: 16632-16642