Biorhem. Physiol. Pflanzen 179, 129-143 (1984) LAMMA-Ionenspektren der Fangschleime carnivorer Pflanzen-) GEORG HEINRICH Institut fiir Allgemeine Botanik der Universitat Hamburg, BRD LAMMA Ion Analysis of the Trapping Slimes of Carnivorous Plants Key Term Index: LAMMAion spectra, trapping slimes, carnivorous plants Summary Fingerprints of anions and cations in the trapping slimes of seven Drosera species, of Drosophyllum lusitanicum, three Pinguicula species as well as the pitcher fluid of Nepenthes alata have been analysed by a laser microprobe mass analyzer (LAMMA). All trapping slimes contain the cations that have been found in the Nepenthes pitcher fluid although in much smaller concentrations. Divalent cations as Mg and Ca prevail, whereas K and especially Na occur in smaller amounts. In the spectra of anions Cl predominates. Zusammenfassung Kationen- und Anionen-Fingerprints der Fangschleime von sieben Drosera-Arten, von Droso- phyllum lusitanicum, drei Pinguicula-Arten und des Kannensaftes von Nepenthes alata wurden mit Hilfe des Laser-Microsonden-Massen-Analysators (LAMMA) angefertigt. In allen Fangschleimen lie- gen die gleichen Kationen wie im Kannensaft von Nepenthes-Arten vor, allerdings in weit geringerer Menge. Hiiufig dominieren die divalenten Kationen Mg und Ca, wahrend K etwas und Na noch star- ker zuriicktritt. Im Anionenspektrum iiberwiegt Cl. Einleitung Hochmoorpflanzen miissen mit geringcn Nahrsalzgchalten vorliebnehmen (KUSEL- FETZMAl'\N 1982). Im ombrogenen Wasser verschiedener Niedermoorgebiete der BRD er- rcichen dieMg- und Ca-Wcrte nur in seltenen Fallen 2 mg, die K-Werte 1 mg/I. Die in Kustennaho auftretenden hohen Na-Gehalte (bis 9 mg/I) nehmen mit zunehmcnder Entfernung zum Meer kontinuierlich ab (MULLER 1973). Der Mineralstoffgehalt der Hochmoorpflanzcn ist, dem Angebot entsprechend, ebenfalls gering (MALMER und SJORS 1955), was besonders der Vergleich mit Pflanzen anderer Standorte deutlich macht (BAl:MEISTER und ERNST 1978). Der Aschegehalt von Sphagnum cuspidatum erreicht nicht 2 %, wahrend er bei SiiBgrasern meist 7 % der Trockensubstanz, bei Brennesseln uber 13 %und bei der Wasserpest Elodea fast 18 % ausmacht. Einige Hochmoorpflanzcn, die keine Mykorrhiza besitzen, verschaffen sich durch Tierfang zusatzliche Nahrung. Mehrere carnivore Pflanzen scheiden einen Fangschleim aus, mit dessen Hilfe hauptsachlich Insekten gefangen werden. lonen liegen im Fang- 1) Herrn Prof. Dr. OTTO HARTEL, Graz, nachtraglieh zum 70. Geburtstag gewidmet 9 Biochcm. PhysioI. Pflanzen, Ed. 179
Transcript
Biorhem. Physiol. Pflanzen 179, 129-143 (1984)
LAMMA-Ionenspektren der Fangschleime carnivorer Pflanzen-)
GEORG HEINRICH
Institut fiir Allgemeine Botanik der Universitat Hamburg, BRD
LAMMA Ion Analysis of the Trapping Slimes of Carnivorous Plants
Key Term Index: LAMMA ion spectra, trapping slimes, carnivorous plants
Summary
Fingerprints of anions and cations in the trapping slimes of seven Drosera species, of Drosophyllumlusitanicum, three Pinguicula species as well as the pitcher fluid of Nepenthes alata have been analysedby a laser microprobe mass analyzer (LAMMA). All trapping slimes contain the cations that havebeen found in the Nepenthes pitcher fluid although in much smaller concentrations. Divalent cationsas Mg and Ca prevail, whereas K and especially Na occur in smaller amounts. In the spectra of anionsCl predominates.
Zusammenfassung
Kationen- und Anionen-Fingerprints der Fangschleime von sieben Drosera-Arten, von Drosophyllum lusitanicum, drei Pinguicula-Arten und des Kannensaftes von Nepenthes alata wurden mitHilfe des Laser-Microsonden-Massen-Analysators (LAMMA) angefertigt. In allen Fangschleimen liegen die gleichen Kationen wie im Kannensaft von Nepenthes-Arten vor, allerdings in weit geringererMenge. Hiiufig dominieren die divalenten Kationen Mg und Ca, wahrend K etwas und Na noch starker zuriicktritt. Im Anionenspektrum iiberwiegt Cl.
Einleitung
Hochmoorpflanzen miissen mit geringcn Nahrsalzgchalten vorliebnehmen (KUSELFETZMAl'\N 1982). Im ombrogenen Wasser verschiedener Niedermoorgebiete der BRD errcichen dieMg- und Ca-Wcrte nur in seltenen Fallen 2 mg, die K-Werte 1 mg/I. Die inKustennaho auftretenden hohen Na-Gehalte (bis 9 mg/I) nehmen mit zunehmcnderEntfernung zum Meer kontinuierlich ab (MULLER 1973). Der Mineralstoffgehalt derHochmoorpflanzcn ist, dem Angebot entsprechend, ebenfalls gering (MALMER und SJORS1955), was besonders der Vergleich mit Pflanzen anderer Standorte deutlich macht(BAl:MEISTER und ERNST 1978). Der Aschegehalt von Sphagnum cuspidatum erreichtnicht 2%, wahrend er bei SiiBgrasern meist 7% der Trockensubstanz, bei Brennesselnuber 13 %und bei der Wasserpest Elodea fast 18 %ausmacht.
Einige Hochmoorpflanzcn, die keine Mykorrhiza besitzen, verschaffen sich durchTierfang zusatzliche Nahrung. Mehrere carnivore Pflanzen scheiden einen Fangschleimaus, mit dessen Hilfe hauptsachlich Insekten gefangen werden. lonen liegen im Fang-
1) Herrn Prof. Dr. OTTO HARTEL, Graz, nachtraglieh zum 70. Geburtstag gewidmet
9 Biochcm. PhysioI. Pflanzen, Ed. 179
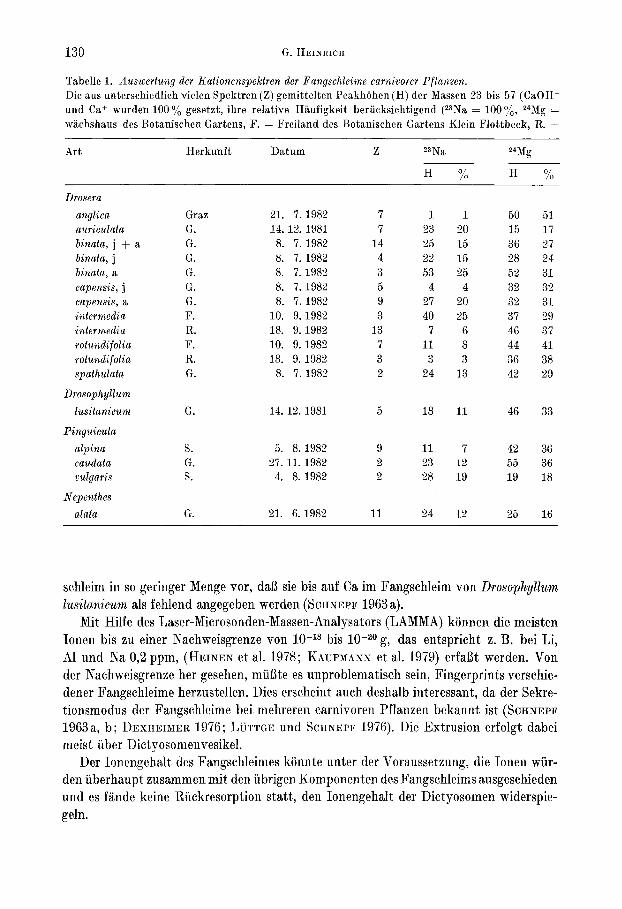
130 G. HEINRICH
Tabelle 1. Auswertung der Koiionenspekiren der Fangschleime carnivorer Pflanzen.Die aus unterschiedlich vielen Spektren (Z) gemittelten PeakhOhen (H) der Massen 23 bis 57 (CaOH+und Ca+ wurden 100% gesetzt, ihre relative Haufigkeit beriicksichtigend (23Na = 100%, 24Mg =wachshaus des Botanischen Gartens, F. = Freiland des Botanischen Gartens Klein Flottbeck, R. =
Art Herkunft Datum Z 23Na 24Mg
H % H %
Drosera
anglica Graz 21. 7.1982 7 1 1 50 51auriculata G. 14. 12. 1981 7 23 20 15 17binata, j + a G. 8. 7. 1982 14 25 15 36 27binata, j G. 8. 7. 1982 4 22 15 28 24binata, a G. 8. 7. 1982 3 53 25 52 31capensis, j G. 8. 7. 1982 5 4 4 32 32capensis, a G. 8. 7. 1982 9 27 20 32 31intermedia F. 10. 9. 1982 3 40 25 37 29intermedia R. 18. 9. 1982 13 7 6 46 37rotundifolia F. 10. 9. 1982 7 11 8 44 41rotundifolia R. 18. 9. 1982 3 3 3 36 38spathulata G. 8. 7. 1982 2 24 13 42 29
Drosophyllum
lusitanicum G. 14. 12. 1981 5 18 11 46 33
Pinguicula
alpina S. 5. 8.1982 9 11 7 42 36caudata G. 27.11.1982 2 23 12 55 36vulgaris S. 4. 8.1982 2 28 19 19 18
Nepenthes
alata G. 21. 6. 1982 11 24 12 25 16
schleim in so geringer Menge vor, daB sie bis auf Ca im Fangschleim von Drosophyllumlusitanicum als fehlend angegeben werden (SCHNEPF 1963a).
Mit Hilfe des Laser-Microsonden-Massen-Analysators (LAMMA) konnen die meistenIonen bis zu einer Nachweisgrenze von 10-18 bis 10-20 g, das entspricht z, B. bei Li,Al und Na 0,2 ppm, (HEINEN et al. 1978; KAUFMANN et al. 1979) erfaBt werden. Vonder Nachweisgrenze her gesehen, mufite es unproblematisch sein, Fingerprints verschiedener Fangschleime herzustellen. Dies erscheint auch deshalb interessant, da der Sekretionsmodus der Fangschleime bei mehreren carnivoren Pflanzen bekannt ist (SCHNEPF1963a, b; DEXHEIMER 1976; LUTTGE und SCHNEPF 1976). Die Extrusion erfolgt dabeimeist iiber Dictyosomenvesikel.
Der Ionengehalt des Fangschleimes konnte unter der Voraussetzung, die Ionen wiirden iiberhaupt zusammen mit den iibrigen Komponenten des Fangschleims ausgeschiedenund es Iande keine Riickresorption statt, den Ionengehalt der Dictyosomen widerspiegeln.
Ionenspektrum von Carnivoren-Fangschleimen 131
und etwas Mg0 2H+) sind aufgelistet. Die Peakhiihen von Na t und der Hauptmassen von Mg+, K'79%, 39K = 93%, 40Ca = 97%) korrigiert und in % der Peakintensitaten angegeben. G. = GeRaakmoor, S. = Silvretta; a = alt, j = jung
Versuchspflanzen sind Drosera (D.) auriculaia BACKH. ex PLANCHON, D. binata LABILL., D.spathulata LABILL., D. capensis L. aus dem Gewachshaus des Botanischen Gartens Hamburg, KleinFlottbek; ferner D. intermedia HAYNE und D. rotundifolia L. (beide aus Bedakesa, nordlich vonBremerhaven [BRD] stammend) aus dem Moorgrund des Botanischen Gartens und vom Naturstand-ort Raakmoor, Hamburg 62, und D. anglica HUDSON aus dem Botanischen Garten der UniversitatGraz. AuBerdem wurden Drosophyllum lusitanicum (L.) LINK, Pinguicula (P.) caudata SCHLECHTEND.aus dem Gewachshaus des Botanischen Gartens Klein Flottbek, P. alpina L. und P. vulgaris L. ausdem Fimbertal, Silvretta, untersucht. Vergleichsanalysen wurden mit dem Kannensekret von Ne-pentes alata durchgefiihrt.
Von den genannten Pflanzen wurde jeweils cine geringe Menge des Fangschleims auf 3 mmgroBe befilmte Gold- oder Kupfernetze (D. anglica) fiir die Elektronenmikroskopie mit Hilfe einervergoldeten Pinzette aufgenommen. Jeweils 2 Netze wurden gefriergetrocknet, urn eine mikrobielleUmsetzung und Veranderungen der Probe infolge eventuell eintretender Kristallisationsprozesse zuverhindern, mehrere andere Netze wurden jeweils luftgetrocknet. Die Messung erfolgte meist inner-halb von 2 Wochen mit dem Laser-Microsonden-MassenAnalysator (LAMMA®) der Fa. Leybold-Heraeus. Dabei werden die Proben durch ein Liehtmikrosknp betrachtet. Ein Pilot-Laser (He-Ne)kennzeichnet die Stelle, an der mit Hilfe eines Hochleistungslasers (Yag-Nd) ein Loch in das Praparat
9'
132 G. HEINRICH
geschossen wird (HEINEN et al. 1978; KAUFMANN et al. 1979). Die durch den Laserimpuls aus demProbenvolumen von etwa 10-13 cm3 gebildeten positiven bzw. negativen lonen werden in einenFlugzeit-Massenspektrometer analysiert.
Je Anregungsimpuls kann ein komplettes Massenspektrum der positiven bzw. negativen lonenaufgezeichnet werden. Radikale werden ihrem Radikalcharakter nach gekennzeichnet, die Kennzeichnung der Ionen nach ihrer Wertigkeit wird meist unterlassen.
Ergebnisse
Die Gehalte der divalenten Kationen Mg und Ca uberwiegen in den meisten LAMMASpektren der Fangschleime. In Tabelle 1 sind die aus bis 14 Einzelspektren gemitteltenPeakhohen fur Na+, Mg+, K+ und Ca+ aufgefUhrt, zur besseren Vergleichsmoglichkeitist auBerdem die Gesamtintensitat der Hauptmassenpeaks dieser 4 Elemente willkiirlich 100 %gesetzt worden und die jeweiligen Einzelintensitaten in %davon angegeben.Die in den meisten Spektren ebenfalls auftretenden Spuren von AI+, Sit, Mn ! und Fe !
werden bei der Berechnung ausgeklammert. Bei den Elementen Mg, K und Ca wirdjeweils die Hauptmasse herangezogen und eine Korrektur in Berucksichtigung der relativen Haufigkeit des jeweiligen Isotops durchgefUhrt. Diese Komplikationen entfallenbei den Reinelementen 23Naund 27AI. Die Peakhohen fUr 25Mg, 26Mg, 27AI, 41 K und 55Mn
und ein Peak 57, der hauptsachlich CaOH+ reprasentiert, werden in Tabelle 1 zusatzlich aufgefUhrt. Wahrend die Masse 40 mit 40Ca gleichgesetzt werden kann (4°K hateine relative Haufigkeit von 0,02, 40Ar kommt nicht in Frage), so stort bei 41K das Kation MgOH+, bei 43Ca kommt AIO+ dazu, bei 44Ca das isobare Ion AIOH+ und SiO+,die in diesem Fall aber nur Bruchteile ansmachen konnen, weil die entsprechendenElemente in sehr geringer Menge vertreten sind. Die Masse 55 entspricht Mn'', bei56Fe storen CaO+ und Mg02+. Der Peak 57 besteht weitgehend aus CaOH+ und Mg02H+.
Eine Aufteilung dieser Peaks auf die Elemente Ca und Mg Iallt schwer. Ebensowenig
Tabelle 2. Ionengehalte im Fangschleim und Kannensaft carnivorer Pflanzen.Blockdiagramm der Ca-, Mg-, K- und Na-Gehalte und die Quotienten Mg/Ca, K/Ca und KINa.j = jung, a = alt, F. = Freiland dcs Botanischen Gartens, R. = Raakmoor
Jonengehalf in %10 20 30 40 50 60 70 80
Co Mg K
90 100 119/ Co fUCa KINO'Drosera anglica t.t 0,04 2
auriculato O/t 0,3 0,8bina/a /-I-a 1 1,1 2o/nata J 0,9 1,2 2,3binata a 7,4 7 0,9capensis j 0,6 0,3 3,2caoensts a 0,8 0.3 O.S/nlermedia F. t.r 0,7 0.8/nlermedia R. 0,9 0.1;- 2,7rotund/folia F 1 0.2 trotund/fol/a R. 0.9 0.'1 So]spatrn)!ala 0.7 0.3 1.1
Abb . 1. Koiionenspektren der Fangschleime von 4 Drosera-Arten.Die Cu t -Peaks bei D. anglica entstammen dem Cu-Nct z.G = Gewachshaus .
.co~..'"
.auS
...u.,.:roau1;;
Drosophyllum lusitanicumG.
Abb. 2. Kal ionenspekiren der Eonqschleime von 4 Drosera-Arten und von Drosophyllum lus ilanicum .F = Freiland des Botanischen Gartens, G = Gewachshaus.
•oZ..,N
.J:. N
0 0ua 01"4 ~-•J:
00ue-ItI.
:.::III.
:.::
•:.::01...•0u0"4
•J:
• 0:l<: 0
~~U... 0>........j.l
Pinguicula vulgarisS.
Nepenthes a\ataG.
Abb.3. Kalionenspeklren von 3 Pinguicula-Arlen und von Nepenthes alala.G = Gowaehshaus, S = Silvretta.
......
I~ ~
24C
,""~,...
H-
24'>
24
26C
2H
2/C
W25
2525
2626
,--
26
:rill:
--
-llli:
lllJ
..:
35C
\-_
3C
;CI-
37C
I-37
CI-
~3
7C
I-36
42C
2H
20
-/C
NO
-42
41~
--4
1
4545
45-
4245
--
48
"l1
k5
9l-5
9
-4
9"-
.
59C
59::J CO
71~
71L
=71
71
....24
0
-2
5'
26::l1
0t'l Z
...;.;
05
,-36
35C
I-a
In37
C\-
1ll
!.:
--
:t:
11)
--4
10
041
...42
...0
0_
_43
0...
4548
0...
4945
InIn
0...
11)
0...
11)
In
r+0
...11
)
C0
...::J
:-n0
059
00
Q.
::J...
....0
00
-71
tD"0
71N
::J
0...
11)
(C
-3
::J-
_.;0
-0
11)
In0
Q.
_.0
0In
-0
....
Ionenspektrum von Carnivoren-Fangschleimen 137
ist zu entscheiden, urn wieviel die Hohe des 4oCa-Peaks durch die in seltenen Fallen auftretenden Massen 96 (Ca20+), 97 (Ca20H+) und die haufigeren Massen 112 und 113, dieals Ca202+ und Ca202H+ gedeutet werden, zunehmen mulste. Verglichen mit Na und Kmulste die Menge an Ca undMg noch groBer sein. Ob die bei Gewachshauspflanzen manchmal anzutreffende Masse 75 einen Massenpeak einer organischen Komponente (z. B.C6Ha+) oder Arsen darstellt, ist fraglich. Die unterschiedlichen Ionengehalte der Fangschleime junger und alterer Pflanzen von D. binata und D. capensis (Tabellen 1 und 2)konnten wohl bestehen, allerdings reicht die Zahl der Analysen zu einer statistischenAbsicherung nicht aus.
Tabelle 2 ermoglicht durch die Verwendung von Blockdiagrammen eine schnelleUbersicht iiber die vorliegenden relativen Intensitaten an Na, Mg, K und Ca im Fangschleim der untersuchten Carnivoren. In 10 von 17 gepriiften Fangschleimen reprasentiert Ca mehr als 40%des Kationengehalts, Mg mehr als 30, K mehr als 15 und Na mehrals 10. Ca und Mg zusammen machen bei allen Fangschleimen iiber 50% der Kationenaus, bei der Halite der Fangschleime iiber 70%. Der durchschnittliche Quotient betragtfiir Mg/Ca 0,9, fiir K/Ca 0,5 und fur KINa 2,0.
In Abb. 1, 2 und 3 sind charakteristische Kationenspektren der untersuchten Fangschleime und des Nepenthes- Kannensaftes dargestellt. Abb. 4 und 5 zeigen charakteristische Anionenspektren. Die Fangschleime von D. anglica aus Graz (Abb. 1), D. intermedia (Abb. 2) und D. rotundifolia vom Naturstandort Raakmoor enthalten sehr wenigNa. In den Anionenspektren der hier untersuchten Arten fallen neben zahlreichenaus der Folie und den organischen Substanzen des Fangschleims gebildeten Ionen auchsolche mit der Masse 35 und 37 auf, es handelt sich dabei urn die beiden Cl--Isotope.Die Analysen lassen erkennen, daB die Drosera-Pflanzen aus Graz (Abb.4) und dievom Raakmoor weniger Cl- enthalten als die Pflanzen aus dem Gewachshaus und demFreiland des Botanischen Gartens (Tabellen 1 und 2). Pinguicula-alpina-Blatter sezernieren anfangs viel Cl-, altere Blatter, die Insekten gefangen hatten, zeigten Massonpeaks bei 63 und 79 (Abb. 5), es handelt sich dabei urn P02 - und POa- , die Clv-Peakshaben deutlich abgenommen. Der Anteil an Sulfat- und Nitrationen im Fangschleimvon Blattern, die keine Beute gefangen haben, ist stets sehr gering.
Die im Kannensaft von Nepenthes vorkommenden Ionen sind von mehreren Untersuchungen her gut bekannt (LUTTGE 1964a, b; MORRISSEY 1955; NEMCEK et al. 1966).Urn an diese Untersuchungen ankniipfen zu konnen, wurde hier ebenfalls der aus einergeschlossenen Kanne stammende Kannensaft analysiert. Da der Nepenthes-Kannensaft
Abb. 4. Anionenspektren der Fangschleime von 4 Drosera-Arten.
D. anglica aus Graz und D. rotundifolia vom Raakmoor (R.) weisen kleine Clr-Peaks auf. F = Freiland, G = Gewiichshaus. Die haufig vorkommenden Massen, die teilweise der Folie, weit mehr aberden organischen Komponenten des Fangschleims entstammen, werden folgendermaBen gedeutet:24 = C2- , 25 = C2H-, 26 = C2H2 - , oder CN-, 41 = C2HO-, 42 = C2H20- oder CNO-, 45 = ?,48 = C4- oder 80-, 49 = C4H- oder Na CN-, 59 = CHaCOO-, oder AI02- , 71 = ?, 72 = C6- ,
73 = C6H-. Die Folie enthalt nur wenig Cl-.
f-"
C;:l
00 P ::II 1'1 Z ::0 c:; ~
-25
'26
2'
-- 26
37C
I-35
Ct-
'8
59
717
3
II' 11
5
129
-0 ::;,
1C C o C o o - "0 :J o
(J)
,1---3
7C
I-
5F
759
F:
71
.--'
26C
2H
2-,
CN
u _. ::;,
1C C 0 C 0(J
)0 - "0 -. ::;
,0
2'
C2
25C
,W
63PO
2-
~'~
C,W
IN
aCN
-
co:
'2C
2H2
0-/
CN
O-
>79
P0
3-
Z to "0 to ::;
, -zr IX(;)0 Q -o
17O
W
41C
2H
O
45
59
25C
2W
;.35
CI-
)37
CI-
r------
71
Ionenspektrum von Carnivoren-Fangschleimen 139
einen wesentlich hoheren Ionengehalt aufweist als der Drosera-Fangschleim, fallen imunverdimnten Saft Kristalle aus, die eine sinnvolle Analyse unterbinden. Nach Verdiinnung der Saftes mit dest. H20 im Verhaltnis 1: 10 erhalt man Spektren, in denenmanchmal mehr Na,r als Mg+ vorhanden ist (Abb. 3) oder umgekehrt, dasselbe gilt IurK+ und Ca'. Der Mittelwert aus 11 Spektren weist etwa gleich viel Ca! wie K+ undetwas mehr Mg+ als Na! auf (Tabellen 1 und 2). In den Anionenspektren dominieren diebeiden Cl--Isotope (Abb. 5).
Diskussion
Ein Vergleich der Fangschleimspektren mit Spektren verschiedener Nektare, Milchsafte oder Guttationstropfen aus Hydathoden (HEINRICH, unveroffentlicht) zeigt, daJ3die LAMMA-Fingerprints meist so charakteristisch sind, daJ3 eine Zuordnung zu bcstimmten Sekrettypen moglioh ist. Der Vorteil der LAMMA-Analyse pflanzlicher Exsudate liegt darin, daJ3 mit kleinsten Exsudatmengen ohne groBeren praparativen Aufwand ein Ubersichtsspektrum erhalten wird. Nachteilig ist, daB selbst eine semiquantitative Auswertung der Spektren iiber den Peakhohenvergleich mit gleichmaliig dotiertenFolien bekannter Ionenkonzentration (SCHRODER 1981) schwierig ist und zahlreicheFehlermoglichkeiten beinhaltet. Eine Quantifizierung wird auch dadurch erschwert, daBbestimmte Peaks durch Uberlagerung verschiedener Ionen zustande kommen.
Eine Korrektur jener Elemente, die mehrere Isotope aufweisen, unter Berucksichtigung der relativen Haufigkeit, ist selbst dann nicht unproblematisch, wenn keine isobaren Ionen storen. Die natiirliche Haufigkeit von 24Mg, 25Mg und 26Mg wird mit 78,7;10,13 und 11,17 angegeben (BUDZIKIEWICZ 1980). Als Mittel aus 93 hier durchgefiihrterVersuche ware sie 67,7; 10,09 und 17,26. Dies bedeutet, daB der in Tabelle 1 gewahlteKorrekturfaktor fur 24Mg zu niedrig gewahlt ist.
Gefriergetrocknete und lnftgetrocknete Praparate des Fangschleims von Droseralassen keine signifikanten Unterschiede im Ionengehalt erkennen. Die vorliegendenIonenmengen sind so gering, daB zumindest bei lichtmikroskopischer Priifung keineKristalle zu erkennen sind, die eine sinnvolle Analyse wegen der inhomogenen Probenverteilung verhindern, wie das etwa bei den Kannensaften von Nepenthes der Fall ist.Diese lassen sich auf Grund ihres etwa zehnmal hoheren Ionengehaltes nur nach starkerVerdiinnung und/oder Gefriertrocknung mit dem LAMMA messen.
Die Drosera-Analysen wurden zunachst nur an Gewachshauspflanzen durchgefiihrt.Da Diingung in den benachbarten Gewachshausern zu einer Verfalschung der Ionengehalte in den Fangschleimen fiihrten konnte, wurde der Fangschleim zusatzlich am
Abb. 5. Anionenspekiren des Fangschleims von Drosophyllum lusitanicum, 2 Pinguicula-Arten undNepenthes alata.Junge P.· Blatter weisen starke CI-·Peaks auf, alte mit gefangenen Insekten starke P02-. und P03-
Peaks. Bei Nepenthes fallen die Signale des Untergrunds fast weg, zumal es sich um 1: 10 mit dest.H20 verdiinnten Kannensaft handelt, G = Gcwachshaus, S = Silvretta.
140 G. HEINRICH
naturliehen Standort entnommen. Bei diesen Pflanzen sind die Na- und Cl-Gehaltesehr gering, K tritt ebenfalls zuriick, ihr Mg- und Ca-Anteil ist hoch. Die hohen NaCl
Gehalte in Stralsennahe sind durch das winterliche Salzstreuen bedingt. Dies wird inGraz unterlassen, was die niederen Na t- und Clr-Peaks bei D. anglica, die unmittelbar
neben der StraBe kultiviert wird, bedingt.Dati im Fangschleim Ionen vorliegen miissen, gleichgiiltig, ob diese von der Pflanze
oder von der Beute stammen, ist auch daraus ZI1 folgern, da13 sie als Effektoren der pro
teolytischen Enzyme essentiell sind. Die Kenntnis iiber die Verdauungsenzyme dercarnivoren Pflanzen hat sich seit der Zusammenfassung des Wissens durch SCHMUCKER
und LINNEMANN (1959) wesentlich vermehrt. Allerdings sind nicht die Verdauungsenzyme aller carnivorer Pflanzen so ausgiebig untersucht, wie die aus dem Kannensekret von Nepenihes-i;.uteri (AlI'lAGASE 1972; NAKAYAMA and AMAGASE 1968; AMAGASE
et al. 1969; TOKES et al. 1974) oder die von Dionaea muscipula (ROBINS and JUNIPER
1980). Bei einigen Enzymen ist inzwischen bekannt, welche Ionen die Aktivitat beeinflussen. Nicht nur Phosphorylierungsreaktionen werden durch Mg2+ aktiviert, sondern
auch die als Exopeptidase wirkende Leucinaminopeptidase (SMITH 1951). Diese findetsich im Kannensaft von Nepenthes nach Beutefang, im sterilen Saft fehlt sie ebenso wiebei D. binata(LUTTGE 1964 b). Die autokatalytische Bildung der Endopeptidase Trypsin
aus Trypsinogen wird stark durch Ca2+ beeinfluLlt (SMITH 1951). Die Metalle, die Glycyl-lleucin-dipeptidase aktivieren, sind gewebespezifisch. Im Kaninchenmuskel erfolgt ihreAktivierung durch Mn2+, in der Rattenmuskulatur durch Zn2+. Prolidase, die ebenso wie
die Glycyl-l-leucin-dipeptidase im Saft gefutterter Klappfallen von Dionaea gefundenwurde (LUTTGE 1964 b), wird ebenfalls durch Mn2+ aktiviert.
Ca wird bekanntlich durch den Transpirationsstrom verfrachtet und ist phloemimmobil. Der Siebriihrensaft und alle Organe, die uberwiegend durch den Assimilatestrom beliefert worden, sind dementsprechend calciumarm (ZIEGLER 1963), weisen aberhohe K-Gehalte auf (PEEL and WEATHERLEY 1958; TAMlI'lES and VAN DIE 1964). Beidem Xylemschmarotzer Viscum album ist der prozentuelle K-Anteil gering, der Mg- und
Ca-Anteil dagegen aber hoch, bei den Phloemparasiten Orobranche ramosa, Cuscutaeuropaea und Monotropa hypopitys verhalten sich dagegen die Gehalte der ein- und
zweiwertigen Ionen gerade umgekehrt. Der hohe prozentuale Ca- und Mg-Anteil in den
Fangschleimen der hier untersuchten Arten weist auf eine Xylemversorgung der Ten
takel hin, die ja auch tatsachlich bei den Drosera-Arten gegeben ist. Im Gegensatz
zu den Drosera-Tentakeln, in die nur Xylemelemente fiihren, liegen in den Drosophyllum-Tentakeln auch Phloemelemente. Allerdings weist der Fangschleim von Drosophyllum nicht mehr K als der von Drosera auf. Dies ist auch verstandlich, da die Tantakel,
auf welche Weise auch immer, zusatzlich mit Assimilaten beliefert werden mussen.Das beweist der hohe Polysaccharidgehalt des Fangschleims, dessen Hydrolse beiDrosophyllum Galaktose, Arabinose, Xylose, Rhamnose und Glucuronsaure ergibt(SCHNEPF 1963a).
Der Ionengehalt des Tentakel-PreBsaftes konnte anders sein als der des Fangschleims.
Dies ware nicht unwahrscheinlich, zumal das KiCa-Verhaltnis im Fangschleim durchschnittlich kleiner als 1 ist. Ein derartiges Verhaltnis der loslichen Ionenanteile ist nach
Ionenspektrum von Carnivoren-Fangschleimen 141
HORAK und KINZEL (1971) fUr physiologisch calciotrophe Pflanzen, dazu gehoren z. B.aIle bisher gepriiften Crassulaceen, charakteristisch (KINZEL 1971, 1972, 1982). Wie dieals okologisch calcifug geltende Drosera (SCHMUCKER und LINNE~IANN 1959) mit demwenigen Ca umgeht, das sie aus den Hochmoorstandorten bezieht, ist unbekannt.Pinguicula solI kalkhaltigere Boden besiedeln. Das trifft auch fiir die hier untersuchteP. alpina zu, die einem wasserzugigen Boden des Samnauns entnommen wurde, deraus der Verwitterung dunkelgrauer weicher Schiefer und braunlicher Kalksteine entstanden ist. Im Fangschleim der hier untersuchten Pflanzen liegen dieselben lonen wieim Kannensekret von Nepenthes vor, als weiterer Hinweis daliir, daB die lonen bei derVerdauung eine Rolle spielen.
Eine Diskussion daruber, ob die im Fangschleim vorliegenden lonen das lonenverhaltnis in den Dictyosomenvesikeln widerspiegeln, erscheint verfriiht. Vorerst miiBtenMessungen an Tentakelschnitten vorgenommen werden, wobei die bekannte Schwierigkeit auf tritt, die lonen nicht im VerIauf der Praparation zu verlagern bzw. herauszulosen,
Von der Ionenzusammcnsetzung her erscheint ein Vergleich der Fangschlcimc mitden aus absalzenden Halophyten abgeschiedenen Losungen gestattet, wenn auch dielonenmengen sich vollig unterschiedlich verhalten. Tamarix-aphylla-Driisen scheidcnNa, K, Ca und Mg aus (WAISEL 1961; BERRY und THmIsoN 1967), haufig iiberwiegen diedivalenten Kationen Ca und Mg (BERRY 1970), dies trifft auch fur die Drusen von Plumbago capensis zu (SAKAI 1974). Als Anion findet sich aber HC03 , so daf Ca- und MgCarbonat als Calcit (CaC03) und Nesquehonit (MgC03 • 3 H20) vorliegen, wahrend inden Fangschleimcn, ebenso wie im Kannensaft, das Hauptanion CI ist.
Dank
Dcr Fa. Leybold-Heraeus, Koln (BRD), dort besonders Herrn Dr. H. J. HEINEN, ferner HerrnDr. W. SCHRODER, Jiilich, danke ich fiir die Benutzung der LAMMA-Gerate und fiir ihre Hilfsbereitschaft. Den Gartnern des Botanischen Gartens Hamburg, besondcrs Herrn P. EGGERS, danke ichfiir ihre Hilie bei der Beschaffung der Pflanzen, Herrn BRAEUTIGAM fiir das Umzeichnen der Spektren,Frau C. ADAMI fiir Fotoarbeiten, Herrn Dr. H. NOTHDURFT fiir Pflanzenbestimmung, der DeutschenForschnngsgemeinschaft fiir eine Reisebeihilie.
Literatur
AMAGASE, S.: Digestive enzymes in insectivorous plants. III. Acid proteases in the genus Nepenthesand Droserapellata. J. Biochern. 72, 73-81 (1972).
AMAGASE, S., NAKAYAMA, S., and TSUGITA, A.: Acid protease in Nepenthes. II. Study on the specificity of nepenthesin. J. Biochem. 66, 431-439 (1969).
AMAGASE, S., MORI, M., and NAKAYAMA, S.: Digestive enzymes in inseetivorous plants. IV. Enzymatic digestion of insects by Nepenthes secretion and Droserapeltate extract: proteolytic and chitinolytic activities. J. Biochem, 72, 765-767 (1972).
BAU:'IEISTER, W., und ERNST, W.: JVIineralstoffe und Pflanzenwachstum. 3. Aufl. Fischer, StuttgartNew York 1978.
BEHRY, W. L.: Characteristics of salts secreted by Tamarix aphylla. Amer. J. Bot. 57, 1226-1230(1970).
142 G. HEINRICH
BERRY, W. L., and THOMSON, W. W.: Compositi on of salt secrete d by salt glands of T nmariz aphylla.Canad. J . Bot . 45, 1774-1775 (1972).
BUD ZIKIEWlCZ, H.: Massenspcktro mctric. Ein e Einfiihrung. Studienbiicher der instrument ellen Analytik, (E dit . F AHR , E.). 2. Aufl, Verlag Chemie, Physik Verlag, Weinheim 1980.
CHANDLER, G. E., and ANDE RS ON, J. W.: St udies on th e origin of some hydrolytic enzymes associated with th e leaves and tentacles of Drosera species and th eir role in heterotroph ic nutrit ion. NewPhytol. 77, 51- 62 (1976).
DEXH EDIER, J .: Etude de la secre tion de mucilage par les cellules des glandes digcsti ves de Drosera(D. rotundi(olia L. ; ct D. capensis L.). Applicati on de quelques t echn iqu es cyt ochomiques.Cyt obioL 13,307- 321 (1976).
H EINEN, H. J ., WECHSUN G, R, VOGT, H., HILLENKAMP, F ., und KAUFMANN, R: Laser MikrosondenMassen-Analysa tor LAlIIMA. Biochem. Umscha u 2, 346-354 (1978).
HORAK, 0. , un d KINZEL, H.: Typen des Mineralstoffwcchsels bei hoheren Pflanz en. Ostcrr . Bot . Z.119, 475-495 (1971).
KAunIANN, R., HILLENKA}IP, F., and WECHSUNG, R : The laser microprob e mass analyzer (LAlIIlIIA):A new instrument for biom edical microprobe ana lysis. Med. Pro gr. TechnoL 6, 109- 121 (1979).
KINZEL, H.: Die calcicolen und calcifugen, basiphiJen und acidophilen Pflanzen. Pflanz enokologiaund Mineralstoffwechsel, pp. 216-380. (Edit . KINZEL, H.) Ulmer, Stut tgart 1982.
KUSEL-FETZ}IANN, E.: Hochmoorpflanz en. Pflanz enokolo gie und Mineralstoffwechsel, pp. 411 - 471.(E dit . KINZEL, H.). Ulmer , St ut tgart 1982.
LUTTGE, U.: Untersuchungen zur Physiologic der Carnivoren-Drusen, Ber. Dtsch. Bot. Ges. 77,181-187 (1964a).
LUTTGE, U.: Unte rsuc hu ngen zur Ph ysiologie der Carniv oren-Driisen. 1. Die an den Verdauungsvorgangen bet eiligt en Enz ym e. Planta 63, 103-117 (1964 b).
LUTTGE, U., and SCHNE PF, E.: Organi c Substa nces. Encyclopedia of Plant ph ysiol, New Series, Vol.2, Part B. Tran sport in Plants II. Part B. Tissues and Organs. (Edit . LUTTGE, U., and Pl nIAN,1\1. G.). pp. 244-277. Springer, Berlin-Heidelberg-New York 1976.
MAL~IER, N., and SJORS, H.: Some determinat ions of elementary consti t uents in mire plants andpeat. Bot. Notiser 108, 46-80 (1955).
MORRISSEY, S.: Chlorid e ions in th e secretion of the pitcher plant. Nature 176, 1220-1221 (1955).
MULLER, K: Okologische un d vegeta t ionsgeschichtliehe Untersuchungen an NiedermoorpflanzenSta ndorten des ombrotrophen Moores unter besond erer Beriicksichti gun g seiner Koik e und Seenin NW-D eutschland. Beitr. BioL Pflanz en 49, 147-235 (1973).
NAKAYA1IA, S., and A1IAGASE, S.: Acid protease in Nepenthes: partial purification and propertiesof the enzyme. Pro c, J ap. Acad, 44, 358-362 (1968).
NEMCEK, 0., SIGLER, K. and KLEINZELLER, A.: Ion transport in the pitcher of Nepenthes henryana.Bioehim. Biophys. Acta 126, 73- 80 (1966).
P EEL, A. J. , and WEATIIERLEY, P. E. : Composit ion of sieve- tube sap. Nature 184, 1955-1956(1959).
ROBINS, R J ., and J UNIPER, B. E.: The secre tory cycle of Dionaea muscipula EI,LIS. IV. The enzymology of the secre tion. New Ph ytol. 86, 401-412 (1980).
SAKA I, W. S.: Scanning electron microscopy and energy dispersive X-ra y an alysis of cha lk secre tingleaf glands of Plumbago capensis. Amer. J. Bot. 61, 94-99 (1974).
SCHMUCKER, T. , und LINN EMANN, G.: Carnivorie. Handbuch PflanzenphysioL XI, pp. 198-283.(E dit . RUHLAND, W.) Springer, Berlin, Gott ingen, Heidelberg 1959.
SCHNEPF, E.: Zur Cytologie und Ph ysiologie pflanzli cher Dr iisen. 1. Teil. Uber den Fangschleim derl nsektivoren. Flora 153, 1-22 (1963a).
Ion enspektrnm VOII Carnivoren-Fangschleimen 143
SCHNEPF, K : Zur Cyt ologie und Ph ysiologic pflan zIicher Druse n. 2. Teil. "O ber die Wirku ng vonSauerstoffentzug und von Atrnungsinhibitoren auf die Sekretion des Fangschleims von Drosophyllul/I und auf die Feinst rukt ur der Dru sonzellen. Flora Hi3, 23- 48 (1963 b).
SCHNEPF, E.: "Ober die Wirkung von Hemmst offen der Proteinsynthes e auf die Sekre tion des Kohlenhydrat- Fangschleimes VO II Drosophyllumlusitanicum. Planta 103, 334- 339 (1972).
SCIIRODER, W. H.: Quantitative LAMMA ana lysis of biological specimens. I. Sta ndards. II. Isotopelabeling. Fresenius Z. Anal. Chern. 308, 212-217 (1981).
SMITH, K L.: Proteolytic En zymes. The enzymes. Chemistry and mechanism of action. Vol. 1.Par t 2, pp. 793- 872. (E dit. SU:MNER, J. B., and MY RB ACK, K.) Academic Pr ess, New York 1951.
TA:\IMES, P.:\1. L., and VAN DIE, J.: St udies on phlo em exudation from Y ucca flaccida HAW. I. Someobserv at ions on th e ph enomenon of bleeding and th e composition of the exudate . Acta Bot . Neer!.13, 76-83 (1964).
TOKES, Z. A., Woox, W. C., and CHA~!BER , S. ~I. : Digestive enzymes secreted by th e carn ivorousplant Nepenthes macfarlanei L. Planta 119, 39-46 (1974).
WAISEL, Y. : Ecological studies on Tn marix aphylla (L.) KARST. III. The salt economy. Plant andSoil 13, 356- 364 (1961).
ZIEGLER, H.: Der Ferntransport organischer Stoffe in den Pflanz en. Naturwiss. 60, 177-186 (1963).
Eingegangen am 4. Marz 1983 ; akzeptiert am 16. Juni 1983
Anschrif t des Verfassers: Prof. Dr. GEORGHEINRICH, Institut fur Pflan zenphysiologie, Schubertst ratle 51, A - 8010 Graz, Osterreich.