62
c 2010 W. Skrandies, Aulweg 129, D-35392 Giessen http://geb.uni-giessen.de/geb/volltexte/2008/6504/
| Date post: | 04-Jun-2018 |
| Category: |
Documents |
| Upload: | trinhkhuong |
| View: | 226 times |
| Download: | 0 times |

KOGNITIVE
NEUROPHYSIOLOGIE DES
MENSCHEN
HUMAN COGNITIVE
NEUROPHYSIOLOGY
c© 2010 W. Skrandies, Aulweg 129, D-35392 Giessenhttp://geb.uni-giessen.de/geb/volltexte/2008/6504/

ImpressumHerausgeber: Wolfgang Skrandies
c© 2010 W. Skrandies, Aulweg 129, D-35392 [email protected]
Editorial Board:M. Doppelmayr, SalzburgA. Fallgatter, WürzburgT. Koenig, BernH. Witte, Jena
ISSN 1867-576X
ii Human Cognitive Neurophysiology 2010, 3 (1)

Kognitive Neurophysiologie des Menschen wurde im Jahr 2008 gegründet. Hier sollenwissenschaftliche Artikel zu Themen der kognitiven Neurophysiologie des Menschen er-scheinen Sowohl Beiträge über Methoden als auch Ergebnisse der Grundlagen- und klinischenForschung werden akzeptiert. Jedes Manuskript wird von 3 unabhängigen Gutachtern beurteiltund so rasch wie möglich publiziert werden.Die Zeitschrift ist ein elektronisches ”Open Access”-Journal, ohne kommerzielle Interessen;http://geb.uni-giessen.de/geb/volltexte/2008/6504/.
Eine dauerhafte Präsenz der Zeitschrift im Internet wird durch die Universität Giessengewährleistet.
Human Cognitive Neurophysiology was founded in 2008. This journal will publish contribu-tions on methodological advances as well as results from basic and applied research on cogni-tive neurophysiology. Both German and English manuscripts will be accepted. Each manuscriptwill be reviewed by three independent referees.This is an electronic ”Open Access”-Journal with no commercial interest, published athttp://geb.uni-giessen.de/geb/volltexte/2008/6504/.
Online presence is guaranteed by the University of Giessen.
2010, 3 (1) Kognitive Neurophysiologie des Menschen iii

Instructions for Authors
Only original and unpublished work will be considered for publication unless it is explicitly statedthat the topic is a review. All manuscripts will be peer-reviewed. Both German and Englishversions are acceptable. After publication, the copyright will be with the editor of the journal.Usage of published material for review papers will be granted. Manuscripts (as WORD or TEXfiles ) should be sent to [email protected].
Organization of manuscripts: The title page with a concise title should give the authors’ names,address(es), and e-mail address of the corresponding author. The manuscript should includean abstract in English (maximum 300 words). Organize your work in the sections Introduction,Methods, Results, Discussion, and Literature. Please also supply a short list of keywords thatmay help to find your publication.
Illustrations: All figures should be submitted as jpeg or Coreldraw files. Please supplyfigure legends that explain the content of the figures in detail. Since this is an electronic journalcolor figures will be published free-of-charge.
The Literature should only include papers that have been published or accepted for publication.The reference list should be in alphabetical order by author. In the text, references should becited by author(s) and year (e.g. Johnson, Hsiao, & Twombly, 1995; Pascual-Marqui, Michel, &Lehmann, 1994; Zani & Proverbio, 2002).
Examples of reference formatJohnson, K., Hsiao, S., & Twombly, L. (1995). Neural mechanisms of tactile form recognition. In
M. Gazzaniga (Ed.), The Cognitive Neurosciences (p. 253-267). Cambridge, Mass.: MITPress.
Pascual-Marqui, R., Michel, C., & Lehmann, D. (1994). Low resolution electromagnetic tomog-raphy: a new method for localizing electrical activity in the brain. International Journal ofPsychophysiology , 18, 49-65.
Zani, A., & Proverbio, A. (Eds.). (2002). The Cognitive Electrophysiology of Mind and Brain.San Diego: Elsevier.
iv Human Cognitive Neurophysiology 2010, 3 (1)

EditorialThis issue contains a long review paper by Pierre Etevenon on modified states of conscious-ness. Although this contribution is not a data paper, the reviewers felt that it is of general interestbecause of the subject areas – consciousness and meditation – that are controversially dis-cussed in scientific publications, and because Dr. Etevenon has contributed to this discussionalmost from its beginning.I believe this summary is of interest to the readers. Of course such a personally flavoredoverview should be considered as a supplement to data papers published in the journal.
Wolfgang Skrandies
2010, 3 (1) Kognitive Neurophysiologie des Menschen v

Inhalt — Contents
Inhalt — Contents
P. Etevenon — Meditation as a State of Consciousness: A Personal Account . . . . . 1K. Suzuki & H. Shinoda — Error-related Components and Impulsivity related to Speed
and to Accuracy Trade-off . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 26W. Skrandies — Abstracts of the 18th German EEG/EP Mapping Meeting . . . . . . . 37Announcements — Ankündigungen . . . . . . . . . . . . . . . . . . . . . . . . . . . . 56
vi Human Cognitive Neurophysiology 2010, 3 (1)

P. Etevenon — ”Modified States of Consciousness” revisited
Abstract
P. Etevenon (Hermanville sur Mer, France) — Meditation as a State of Consciousness: A PersonalAccountInsofar as ”consciousness” is considered to be an axiom, different definitions of ”states of consciousness”
can be envisaged. It was in Princeton and later in Paris in Pierre Deniker’s psychiatric clinic that the
author first studied ”pathological states of consciousness” and ”natural states of consciousness”, together
with quantitative EEG analyses of psychiatric and neurological conditions, as well as the effects of hallu-
cinogenic and psychodysleptic drugs. In his opinion, ”states of meditation” and a wider category that he
called in 1972 ”voluntary controlled states of consciousness” are distinct both from these ”altered patho-
logical states” and from the ”natural states of consciousness” typical of wakefulness, sleep and dreams
(particularly in paradoxical or REM sleep). In the Sainte-Anne hospital in Paris the first EEG recordings
of the Zen master Taisen Deshimaru Roshi and his disciples were carried out and EEG spectral analy-
ses computed. In two previous articles (1972 and 1973), the author validated by spectral analysis the
high amplitude hypovariable alpha rhythm discovered earlier by Japanese scientists in their initial EEG
explorations of zazen meditation. Throughout the past ten years, particularly in the United States, neu-
roscientists, continuing the work of Francisco Varela and Antoine Lutz, have made significant progress
in studying states of meditation, mainly with respect to Buddhist practices. This review presents the au-
thor’s own point of view, and summarizes and discusses these recent findings from the perspective of his
ongoing experience of thirty years of research, in which he argues that states of meditation are neither
hallucinations nor sleep stages.
Keywords: Meditation, Consciousness, Electroencephalography, Brain mapping, Awareness, Wakeful-
ness, Sleep stages, REM sleep, Dreams, Yoga, Buddhism, Zen
P. Etevenon — Meditation asa State of Consciousness: A
Personal Account
Pierre Etevenon, DSc, PhD, Directeur deRecherche honoraire INSERM, 71 rue des
verts prés, 14880, Hermanville sur Mer,France
Introduction
Conscience, Consciousness and States ofConsciousness
According to Coxeter (1974), ”the importantprinciple [is] that any definition of a word mustinevitably involve other words, which requirefurther definitions. The only way to avoid avicious circle (Synge, 1951) is to regard cer-tain primitive concepts as being so simple andobvious that we agree to leave them unde-
2010, 3 (1) Kognitive Neurophysiologie des Menschen 1

P. Etevenon — ”Modified States of Consciousness” revisited
fined.” If we want to avoid the kind of circulardefinitions that are to be found in dictionaries,we have to take into account those primitivestatements called axioms. For ”The Man WhoSaved Geometry” (Roberts, 2006), the cham-pion of n-dimensional geometry and builder ofpolytopes, a ”point” was an axiom allowing fur-ther definitions of position, place, and exten-sion as well as of line, plane, space, etc. Inagreement with this great man, a materialist, Iwould, for my part, transpose his concept andargue that ”consciousness is an axiom.”
That basic primary concept enables us todefine and construct different ”states of con-sciousness.” This point of view is not farremoved from that of William James (1902),who, going beyond his empirical attitude,considered (in 1892) that consciousness is aconstantly-changing flux: ”When I say every’state’ or ’thought’ is part of a personal con-sciousness, ’personal consciousness’ is oneof the terms in question. Its meaning we knowso long as no one asks us to define it, butto give an accurate account of it is the mostdifficult of philosophic tasks.” This again isbecause consciousness is an axiom, a basicprimary concept from which different points ofview emerge among visionaries, seekers oftruth, philosophers and scientists, both in thepast and in contemporary society.
Conscience and consciousness refer to twodifferent concepts which were discussed byRoland Fischer in 1975: ”As the Oxford En-glish Dictionary points out, the English lan-guage adapted conscience, that is the ‘privityof knowledge’, from the Latin; so did the Ro-mance languages. But the French, the Ital-ian and the Spanish have but one word forboth conscience and consciousness and only
the context can assist in the differentiation ofthe meaning.” Engelberg in 1972 consideredthat in German (as Hegel, Schopenhauer, Ni-etzsche, and Freud noted): ”Conscience isGewissen and consciousness is Bewusstsein”.Roland Fischer recalled that around 1000 A.D.the monk of St. Gallen, Notker ”teutonicus”,used gewissen as a translation of the Latinconscius – ”so that Conscience originally alsomeant consciousness in the religious-moralsense” – and went on to say that ”the Englishand German languages, at least in commonusage, sometimes behave as if they had for-gotten the near identity of the two words whichonce in fact existed”. Fischer points out that”in English ‘conscious’ as meaning ‘inwardlysensible or aware’ appears first in 1620, ‘con-sciousness’ in 1620 or ‘the state of being con-scious’ in 1678, and ‘self-consciousness’ or‘consciousness of one’s own thoughts’ in 1690.It is also interesting that, according to Whyte(1959), ‘conscious’ whose Latin source hadmeant ‘to know with’ (to share knowledge withanother) now came to mean ‘to know in one-self, alone’.”
In another article, Fischer (1972) analyzedthe difference between what he called objec-tive ‘I’-consciousness and levels of subjective‘Self’-awareness: ”We reserve the concept of‘consciousness’ for a state of mind associatedwith the most intense objective (rational) ‘I’-awareness ‘out there.’ Each consecutive layerof self-awareness with diminishing objectivity‘out there’ is accompanied by an increase insubjective ‘Self’ - Awareness, experienced asmeaning ‘in here’.”
For the French I.T. scientist Anceau (1999), theword conscience covers many notions and hastwo principal forms. In the first of these, it is a
2 Human Cognitive Neurophysiology 2010, 3 (1)

P. Etevenon — ”Modified States of Consciousness” revisited
”state of arousal, of vigilance” which is mani-fested only during wakefulness and which dis-appears during sleep and in comas. The sec-ond form is the ”epitome of superior psychicstates”, a characteristic peculiar to living crea-tures, and beyond that, to the whole universe –indeed, in some cases this form of conscienceis placed in the same category as the divine.According to Anceau, although these two defi-nitions may coexist, the second is spiritual; hethen focuses on the first notion, which encom-passes different forms of conscience imply-ing or not reflection, memory, self-observation,thinking about thinking, feelings, etc. Anceau,who is an expert in parallel computing andthe architecture of micro-processors, consid-ers three main pathways that research intoconscience can follow. The first is dualist, fol-lowing Descartes (Eccles, 1989). The second,which is his own focus, is ”materialist, phys-icalist, methodological (and scientific)” and isrepresented by Crick (1994), Edelman (1992),Dennett (1991), and in France by Changeux(1983), Dehaene et al. (2006) and Naccache(2006). (See also, more recently, La con-science 2008, and ” Consciences et Neuropsy-chologie 2010 ”). The third current of opinionsees conscience as an extension of physicsat the level of quantum mechanics (Penrose,1994): behind the neuronal mechanisms lies aquantum hypercomputer made up of the neu-ronal cytoskeleton which is responsible for theprocess of conscience.
After this general introduction, let me now statemy own point of view. To my mind, conscienceis an axiom – more than that, indeed, for Iwould say that it is primordial in the meaningthat Indian culture attaches to the word, with-out implying any spiritual or religious dimen-
sion (Etevenon, 1974; Etevenon & Santerre,2006). I agree with Spinoza, who makes noseparation between the body and the mind orspirit – my position is thus ‘gnostic’ in JohnScotus Erigena’s sense; the latter’s work hasbeen looked at afresh by two mathematicianswho were invited by Roland Fischer in 1967 fora meeting on ”Interdisciplinary Perspectives ofTime.” There, Günther (1967) and von Foer-ster (Günther & von Foerster, 1967) describedwhat they called ”the logical structure of evolu-tion and emanation” based on a new multivari-ate logic applied to the description and evolu-tion of the universe. They argued that the uni-verse can be looked upon as a sine wave os-cillating around the median line of a quadrantin which the two orthogonal axes are (i) ”ema-nation” (a word used by John Scotus Erigena,810-877 A.D., who considered the universe tobe a theophany, i.e. a revelation of God duringthe process of time) for the horizontal axis – akind of supreme ”determinism” – and (ii) ”evo-lution” for the vertical axis – a ”randomness”that makes for complete freedom in the ge-netic factory which is basically that describedby Jacques Monod (1973). This is a ”truemonism” – a gnostic philosophical point of viewthat brings together both dualism and spiritu-alism with the materialist statements that arealways mutually antagonistic. This is a majorconcept in Indian philosophy (Zimmer, 1953),which we find also in the writings of Sri Au-robindo (1949, 1970) and Jean Gebser (1949,1985), who spoke of an ”integral conscious-ness” and visited the Sri Aurobindo ashram inPondicherry.
2010, 3 (1) Kognitive Neurophysiologie des Menschen 3

P. Etevenon — ”Modified States of Consciousness” revisited
A History of States ofConsciousness and Research onMeditation
The three main categories of states ofconsciousness
In 1965-1966, I was involved in post-doctoralresearch for NATO and thereafter worked as aresearch assistant. It was in Princeton that Ifirst studied ” pathological states of conscious-ness” in animals and in human beings viathe application of quantitative EEG analysesin researching, at the New Jersey Neuropsy-chiatric Institute (NJNPI), the effects of hallu-cinogenic and psychedelic drugs. The Insti-tute was headed by Humphrey Osmond, thefather of psychedelics, and I worked in the neu-ropharmacology department under the lead-ership of Leonide Goldstein, a pioneer andspecialist in quantitative EEG. In Princeton, Istudied the effects of intermittent light stim-ulation on the cortical visual area and sub-cortical visual thalamic nuclei of the rabbit.Specific lateralization of the electrophysiologi-cal responses and their variation after adminis-tration of LSD and amphetamine were studied.At doses producing hallucinations, these drugsinduced hyperarousal and suppressed the pre-viously observed ”cerebral dominance”. ThisLSD facilitation was antagonized by pentobar-bital. (Etevenon, 1967; Etevenon & Boissier,1972). I participated also (Etevenon, 1984)in experiments on the behavioral and EEG ef-fects of psychedelic drugs on human beings.This was the period in which a great deal of re-search was being carried out on LSD and ”al-tered states of consciousness” (Tart, 1969). Ishall not go into any more detail about this do-main of research, which remains nonetheless
a major theme of contemporary studies in theneurosciences and in medicine generally.
From 1967 to 1977, in Paris, I was a neuro-biological scientist with INSERM (Institut Na-tional de la Santé et de la Recherche Médi-cale, the French equivalent of NIH) in charge ofthe ”Experimental neuropharmacological labo-ratory” of Research Unit U-320 INSERM, un-der the direction of J. R. Boissier. We pro-posed (Etevenon & Boissier, 1974) a theo-retical model of schizophrenia which was ananticipation of the later model (Mahowald &Schenck, 1992) of dissociated mixed statesof wakefulness and sleep. At the same timeand for a further period, I studied the effectsof psychotropic compounds on psychiatric andneurological patients in Pierre Deniker’s psy-chiatric clinic in the Centre Hospitalier Sainte-Anne; there, from 1977 until 1986, I was incharge of the ”Quantitative EEG laboratory”,part of the ”Laboratoire d’E.E.G.” of the Sainte-Anne hospital directed by Georges Verdeaux.
During these years I specialized in the quanti-tative EEG (Etevenon, 1985) study of ”patho-logical states” in human beings and of ”natural states of consciousness”, i.e. thedifferent states of wakefulness and arousal,together with sleep stages and paradoxi-cal sleep (REM). I studied, for example, theeffects of the active compound of hashish(Deniker et al., 1974; Etevenon, 1978.) Wecompared and by cluster multivariate analysis(Etevenon, Pidoux & Rioux, 1979) were able todiscriminate between quantified EEG record-ings of two groups of schizophrenic patients(two subgroups: hebephrenics and paranoidschizophrenics, Etevenon et al., 1979) andtwo samples of paired control subjects (twosubgroups: high-alpha and low-alpha sub-
4 Human Cognitive Neurophysiology 2010, 3 (1)

P. Etevenon — ”Modified States of Consciousness” revisited
jects). After initiating in France EEG mappingswith multiple electrodes (Etevenon et al.,1985; Walter et al., 1984), this procedure wasapplied to the EEG mapping of states of wake-fulness under visual (Etevenon et al., 1985)or mental calculation tasks (Etevenon, 1986)and stages of sleep (Etevenon & Guillou,1986). This enabled us to classify stages ofwakefulness and sleep by means of increasedEEG averaged amplitudes (square roots ofEEG power values over posterior occipital andparieto-occipital areas): the minimum EEGamplitude was the desynchronized and acti-vated EEGs recorded in ”active wakefulness,eyes open” under a diffuse vigilance statefollowed by ”quiet wakefulness, eyes closed”with high alpha activity amplitude (definedas ≥ 40µV ‘root-mean-square’ over parieto-occipital and/or occipital areas, characteristicof ”high-alpha” subjects). Between these twophases of ”external wakefulness” we observedthe two stages of what we called ”internalwakefulness”: the mean EEG amplitudes of”stage 1 of drowsiness”, often simultaneouslypresent with subjective hypnagogic imagery,and the mean EEG amplitudes of ”paradoxi-cal sleep (REM)”, frequently associated withdreaming. I went on to make the first dynamicfilm of these EEG mappings from wakefulnessto sleep stages (Etevenon, 1986, 1989), il-lustrating the time flux of dynamic and rapidEEG changes in successive brain mappings.I discovered also that, during REM periodsand after wakening the recorded subjects andinquiring about their dream contents, the EEGmappings and EEG tracings showed selec-tive activated areas related to the subjectivedream contents (Etevenon, 1989). For exam-ple, when an activated left-side area was seen
on the recorded left rolandic EEG location,in the dream content reported after waken-ing the subject was indeed using his or herright hand. However, the observed previousEEG mappings were ”REM mappings” and notsubjective verbal reports, after wakening, ofactual ”dream mappings”. This is so because,in their dream reports, subjects may be car-rying luggage, making a phone call or writingusing their ”right” hand, but the correspondingEEG mappings display only the stimulatedand activated right hand over the contralat-eral ”left” sensory-motor cortical area. ThisEEG activation then produces rapid frequencyactivity with low amplitude desynchronizationover the contralateral ”left” sensory-motor cor-tical area corresponding to an ”active internalwakefulness state” of specific REM activation.
As in the preceding category of pathologicalstates of consciousness, many papers havebeen published on the neuropsychobiology ofnatural states of consciousness, wakefulness,arousal, attention, and mental tasks, as well ason stages of sleep. New articles, reviews andbooks are constantly appearing, with somequite remarkable reports of new findings andnetwork models (Dang-Vu et al., 2008; Hob-son, 2009) in hypnology and oneirology, fol-lowing the studies carried out by Michel Jouvetand his collaborators in Lyon (Jouvet, 2009;Beaubernard, 2009). I shall not for the mo-ment comment on these breakthroughs, whichlie outside the scope of the present review, fo-cusing principally as it does on meditation.
The EEG appears to be a ”marker of statesof vigilance”. ”External wakefulness experi-ences” are linked to environmental stimuliand behavioral responses in the recordedsubject. The activated EEG tracings are
2010, 3 (1) Kognitive Neurophysiologie des Menschen 5

P. Etevenon — ”Modified States of Consciousness” revisited
desynchronized, with small amplitudes andrapid frequency waves. Pfurtscheller’s method– ERD (Event Related Desynchronization) /ERS (Event Related Synchronization) – is wellsuited to the study of successive periods of125 ms accompanied by triggered externalstimuli which may be sensorimotor, auditory(Lebrun et al., 1998, 2001), visual, or evenemotional and/or cognitive complex tasks fol-lowed by recognition and/or choice responses(Pfurtscheller & Aranibar, 1977; Pfurtscheller& Lopes da Silva, 1999). On the other hand,synchronization and relaxation of visual areasinduce high alpha activity similar to scanningalpha waves (Walter, 1953) during ”internalwakefulness experiences” in the course ofattentional tasks performed by the recordedsubject (Ray & Cole, 1985). These inter-nal experiences resemble quiet meditativestates (enstatic) that Roland Fischer called”trophotropic” (Fischer, 1971, 1975, 1976), asopposed to ”ergotropic” meditative states (ec-static). Consequently, it is extremely importantthat groups of normal control subjects be verywell-paired: same age range, right-handed,same sex ratio, balanced high alpha/low alphaor chosen high alpha for greater EEG dynam-ics, same socio-cultural level, etc. Moreover,it is necessary to habituate them to the EEGlaboratory settings, or, depending on theexperimental protocol, to the MEG, MRI orfMRI settings. According to Leonide Goldstein(Goldstein, 1979, 1983; Etevenon, 1986) andto Francisco Varela (Varela, 1995; Varela etal., 2001), it is very important to interview thesubject before, during and after an experimentwith EEG recordings or other brain imagerytechnique, in order to gain access to his or her”inner subjective state of consciousness” in a
neurophenomenological approach combinedwith a more objective research protocol (Lutz,2002).
Since 1963, Indian tradition and culture havebeen of considerable interest to me, especiallyas regards the different methods and practicesof yoga and states of meditation, as well asthe practice of zazen in Japan. This interestprompted me to record with Henrotte (1969,2001) and with Verdeaux in 1972, the EEGs ofa Zen master (Deshimaru, 1981; Deshimaru &Chauchard, 1976) and his disciples and thoseof a Indianist scholar (Lilian Silburn) from theCNRS (cf. the chapter on Lilian Silburn inDescamps, 2006) trained in the Shivaism ofKashmir (Silburn, 1970, 1981-1985); this is avery intense form of meditation that reacheshigh ”peaks of consciousness” (Maslow, 1970)and gives rise to unusual states of conscious-ness such as ecstasy or enstasy (Eliade, 1958,1967). As a consequence of this preliminarypioneering research, I considered ”states ofmeditation” to be part of a larger categoryof states of consciousness which at that time(1972) I called ”voluntary controlled states ofconsciousness”; these are different both from”altered pathological states” and from ”natu-ral states of consciousness” of wakefulness,sleep and dreams (mainly in paradoxical sleep/ REM). As a result of several trips to Indiaand Japan, beginning in 1970, my interestin the inner life and inner experiences (Ma-sui, 1952) was also progressively manifestedover time by the publication of several arti-cles (Etevenon, 1974, 1994, 1999, 2007) andof four books (Etevenon, 1984, 1987, 1990;Etevenon & Santerre, 2006) containing essayson psychotropic drug effects, dreams, yoga,the physiology of wakefulness and sleep, and
6 Human Cognitive Neurophysiology 2010, 3 (1)

P. Etevenon — ”Modified States of Consciousness” revisited
states of consciousness.
The fourth period of my research career, whichbegan in organic chemistry in 1961 (my PhDdates from 1963) but soon drew me towardsneuropharmacology and quantitative EEG(D.Sc. thesis: Etevenon, 1977) in psychiatryand neurology, took place in Caen (Normandy)from 1986 to 1999. There I set up the ”Labo-ratoire de cartographie EEG” (EEG mappinglaboratory) in the ”Centre Esquirol” (the psy-chiatric center headed by the late EdouardZarifian) and later in the INSERM ResearchUnit-320 headed by J.C. Baron (who is nowin Cambridge). There I learned about andapplied new protocols and EEG techniquesin neuropsychology (Etevenon, 1992, 1993;Etevenon et al., 1999), working with normalvolunteers, young subjects who were requiredto carry out complex attentional mental tasksor those requiring auditory selective attention.These protocols were pre-PET scan studiesbecause the INSERM unit U-320 was centeredon a PET scan camera.
The ”modified states of consciousness”(M.S.C.) described in France by by GeorgesLapassade (1987) and later Juan Gonzalezinclude what Tart (1969) calls ”altered statesof consciousness”, such as hypnotic statesand trances, meditation, biofeedback train-ing, etc. The ”voluntary controlled states ofconsciousness” that I described in 1972 canalso be looked upon as a subset of these”modified states of consciousness”; in it areto be found not only ”states of meditation”but also rare states of consciousness suchas the ”shamanic trance” practiced for manycenturies in Mongolia and Siberia. This hasvery recently been studied in EEG mappingwith source localization by Pierre Flor-Henry
(2007) in Canada (as yet unpublished).With this description as a backcloth, I can nowset out a short history of research into medita-tion between 1970 and 2009.
Research into Meditation
Four main bibliographies have been publishedof studies on meditation and consciousness.The first (Timmons & Kamiya, 1970; Tim-mons & Kanellakos, 1974) was drawn up byneuroscientists at the Langley Porter Psychi-atric Institute in San Francisco headed byEnoch Callaway. Joe Kamiya was a pioneerof alpha-wave biofeedback who, together withElmer and Alyce Green (1970) at the Men-ninger Foundation in Topeka, Kansas, initi-ated studies and recordings of yogis. By1970, more than 400 references had alreadybeen reported, beginning mainly in 1957 withthe journal Electroencephalography and Clin-ical Neurophysiology. That review publishedsome seminal papers – by Bagchi and Wenger(1957), who conducted their research on yogaexercises in India; by Kasamatsu, Okuma andTakenaka (1957) describing ”The EEG of ‘Zen’and ‘Yoga’ Practitioners”; and by Das andGastaut (1957) on ”Variations of the electri-cal activity of the brain, heart, and skeletalmuscles during yogic meditation and trance.”However, some years later, in 1973, Gastautstated that the conclusions of his 1957 pa-per written in collaboration with Das were erro-neous because of an earlier misinterpretationbased on unrecognized artifacts. At that time,I published two articles on spectral analysis ofEEGs in the course of zazen posture and yogicmeditation (Henrotte, Etevenon & Verdeaux,1972; Etevenon, Henrotte & Verdeaux, 1973);in 1973, Banquet published his first article on
2010, 3 (1) Kognitive Neurophysiologie des Menschen 7

P. Etevenon — ”Modified States of Consciousness” revisited
transcendental meditation practitioners, whichpre-dated that of Hebert and Lehmann (1977).
In the 1970s, three theoretical articles cameto similar conclusions in presenting two cate-gories of internal states of meditation. Thisis well summarized by Michael Murphy in thefirst chapter of Scientific Studies of Contem-plative Experience: an Overview in the sec-ond major bibliography that I have mentioned(Murphy & Donovan, 1983, first edition); itbrings together 1253 entries dating from 1931to 1983. In it, Murphy writes: ”Roland Fischer,Julian Davidson, and other researchers haveproposed some ways in which internal statesmight be correlated with different physiologicalprofiles (Fischer, 1971; Davidson, 1976)”; hegoes on to say that ”Julian Davidson, RolandFischer, and others have distinguished be-tween two classes of meditation, those that re-lax and those that excite, associating their ef-fects with the trophotropic and ergotropic con-ditions of the central nervous system modeledby Gellhorn and Kiely (Davidson, 1976; Fis-cher, 1971, 1976; Gellhorn and Kiely, 1972)”.That idea remains valid to this day, and echoeswhat I will later in this paper describe as ”medi-tation with objects” and ”meditation without ob-jects.”
A revised and completed edition of the ”Mur-phy and Donovan bibliography” (1999-2009)has since been drawn up by the ”Institute ofNoetic Sciences” in California; this is freelyavailable on the Internet (IONS Bibliographyon Meditation). In their current database, theauthors have recorded 4206 items, covering1937 to 2009. This would appear to be themost extensive bibliography on the subject ofmeditation. Others have been published, suchas that by Thomas Metzinger, with its 140
pages of references of books and articles on”Consciousness - Selected Bibliography 1970-2004” in philosophy, cognitive science, andneuroscience over the last 34 years; essen-tially, this is an attempt to classify the variousdebates that have taken place in English andin German (Metzinger, 1995). This too is freelyavailable on the Internet.
Research in consciousness and into medita-tion began in 1960 (Anand, Chhina & Singh,1961; Wenger & Bagchi, 1961) and devel-oped mainly in the 1970s (Naranjo & Ornstein,1971; Ornstein, 1972) after Maharishi MaheshYogi became, in 1967, a leader among ”NewAge” flower-people and an anti-drug advocate;his early followers were the Beatles, Mia Far-row, and Shirley MacLaine, soon to be fol-lowed by Clint Eastwood, David Lynch and oth-ers. Two names in particular were associatedwith the increasing amount of EEG researchstudies of ”Transcendental Meditation (TM)”,a repeated mantra and inner auditory medita-tion introduced by the Maharishi. These firsttwo associates were Wallace (1970; Wallace& Benson, 1972) and Herbert Benson (1975,1987, 1997; Benson et al., 1982; Benson &Proctor, 1985, 2003; Hoffman, Benson et al.,1982; Lazar et al., 2000, 2007); the latter wasa cardiologist in Harvard and the well-knownauthor in 1975 of a best-seller, The Relax-ation Response, which was followed by severalother books. Their focus of study was the bi-ological and physiological changes occurringduring T.M. and the discovery that a ”wake-ful hypometabolic physiologic state” (Wallace,Benson & Wilson, 1971) was associated with”decreased blood lactate” (Wallace, Benson etal., 1971). Benson is the most quoted authorin the IONS bibliography (97 references), fol-
8 Human Cognitive Neurophysiology 2010, 3 (1)

P. Etevenon — ”Modified States of Consciousness” revisited
lowed by Orme-Johnson (45 references includ-ing Orme-Johnson & Haynes, 1981) and byWallace (23 references). Other scientists havecriticized these pre-1990 results. According toFenwick et al. (1977), for example, ”EEG re-sults showed T.M. to be a method of holdingthe meditators’ level of consciousness at stage‘onset’ sleep”. They found no evidence to sug-gest that TM produced a hypometabolic statebeyond that produced by muscle relaxation,any support for the idea that TM is a fourthstage of consciousness (Murphy & Donovan,1983). A whole series of books containingpapers on TM practice, from its early begin-nings up to the present day, have been pub-lished, mainly in English-speaking countriesand elsewhere in Europe – for example, thoseby Banquet (1973) in France and by Hebertand Lehmann (1977) in Switzerland – as wellas articles from many other countries through-out the world.
From 1970 onwards, research into meditationhas been sponsored in the United States notonly by academic and governmental researchgrants but also with the financial support of pri-vate foundations and institutions such as thevarious T.M. societies and a University ded-icated to the teachings of Maharishi MaheshYogi. Thus it was that in 1967, after ”the publi-cation of Your Maximum Mind, Herbert Bensonlaunched the ‘Mind-Body Medical Institute’, afor-profit research and training initiative in be-havioral medicine, in conjunction with the Dea-coness Hospital in Boston and the HarvardMedical School” (Murphy & Donovan, 1983).
The New Age movement, with personaldevelopment training, multiple successiveconferences and publications, was givensignificant support by the Esalen Institute
(http://www.esalen.org/) founded in Big Sur,California, by Michael Murphy and Dick Price.They were joined in 1962 by Alan Watts andAbraham Maslow, and later by Aldous Huxley,Fritz Perls and early leaders such as ArnoldToynbee, the theologian Paul Tillich, LinusPauling (who was twice awarded the NobelPrize), Carl Rogers, B. F. Skinner, VirginiaSatir, Buckminster Fuller, Timothy Leary andJ.B. Rhine. Since both its founders werestudents of Frederic Spiegelberg, professorof comparative religion and Indic studies inStanford, Esalen soon became a nexus of the1960s counter-culture movement and a centerfor humanistic alternative education with aparticular focus on Indian and Asiatic cultures.As a young man of barely 19 years, MichaelMurphy visited the Sri Aurobindo Ashram inPondicherry and all his later work and re-search was inspired by the ”integral purnayoga” of Sri Aurobindo (Murphy, 1992).
Similarly, the 14th Dalaï-Lama founded the”Mind and Life Institute” in 1990, which, withthe help of the neuroscientist Francisco Varela,developed research, still very much alive, intoBuddhist meditation (Mind and Life, I to VIII,1989, 1990, 1992, 1995, 2000). This researchis at present being followed up in differentneuroscience centers throughout the world byscientists such as Richard Davidson (David-son & Davidson, 1980; Davidson & Harring-ton, 2002) and Antoine Lutz (Lutz et al., 2002,2004, 2007, 2008) who is an associate scien-tist at the Waisman Laboratory for Brain Imag-ing & Behavior at the University of Wisconsin-Madison directed by Davidson.
The major themes of the 4206 items in theIONS bibliography have the following overallfrequency, classified according to key-words:
2010, 3 (1) Kognitive Neurophysiologie des Menschen 9

P. Etevenon — ”Modified States of Consciousness” revisited
Meditation 1696 items; Transcendental medi-tation 539 and ”t.m.” 383; Consciousness 192;Yoga 149; Zen 142; Brain Imagery : EEG 102,MEG 2, MRI 2 including 1 fMRI, TEP 9; Bud-dhist 82 and Buddhism 51; compassion 8 andempathy 17; vipassana 21; kundalini 9. Thislist represents 81% of all of the items in theIONS bibliography items, so that the key-wordscan in themselves be considered significant.I will now focus my attention on the history andresults of the earliest EEG studies of zazenposture meditation. These have given rise tosome interesting findings which have been ver-ified in subsequent studies. I shall then go onto summarize some recent studies of medita-tion in the neurosciences.
Early EEG studies of zazen meditation
Just as Wallace and Benson are renowned fortheir work and writings on transcendental med-itation, two names are associated with studiesof the practice of Zen meditation, and in partic-ular on the ”zazen sitting posture of meditation”(Coupey, 2006) in which subjects sit cross-legged for 30 minutes without moving, in frontof a blank wall, their eyes half-closed lookingtowards the tip of their nose. Akira Kazamatsuinitiated these studies (Kasamatsu, Okuma &Takenaka, 1957; Kasamatsu et al., 1958) andwas followed in 1960 by Tomio Hirai with his”electroencephalographic study of Zen med-itation”, a field of study which he continuedto explore (Hirai, 1974). In their work to-gether (Kasamatsu & Hirai, 1963), they wenton to illustrate their′′electroencephalographicstudy of Zen meditation” (Kasamatsu & Hirai,1966) which proved to be of considerable inter-est. Both were physicians at the University ofTokyo, studying changes in EEG occurring dur-
ing meditation in Zen masters and their disci-ples (48 in all from the Soto and Rinzai centersin Japan); these were divided into 3 groups,based on length of practice, and were com-pared with a control group of 18 subjects and4 older subjects with no experience of medita-tion. Kasamatsu and Hirai made EEG record-ings of 12 channels together with polygraphmeasurements (pulse rates, respiration, gal-vanic skin response), and tested responses tosensory stimuli during meditation. They found(Kasamatsu and Hirai, 1966) that there arefour successive phases in the time-course ofzazen meditation:
Stage 1: Characterized by the appearance ofalpha waves (8-12 Hz) despite eyes being half-open.
Stage 2: Characterized by an increase in theamplitude of persistent alpha waves.
Stage 3: Characterized by a decrease in thefrequency of alpha waves.
Stage 4: Characterized by the appearance ofrhythmical theta trains (7 Hz) at the end of the30 minutes of meditation.
In addition, a ”modulation of the amplitude ofalpha waves” occurred together with the block-ing of theta frequency trains when the medita-tors heard repeated clicks.
I have observed the same results when record-ing a Zen master in the Sainte-Anne hospital inParis (Taisen Deshimaru Roshi) with his disci-ples (Etevenon, Henrotte & Verdeaux, 1973).Later, in his book on Zen and the brain (Deshi-maru & Chauchard, 1976), Deshimaru pro-posed an interesting structural and functional-analysis diagram of zazen and brain neuronalcenters. Paul Chauchard suggested that the”body image” and the ”I” image in the corre-sponding nuclei of brain networks betray re-
10 Human Cognitive Neurophysiology 2010, 3 (1)

P. Etevenon — ”Modified States of Consciousness” revisited
laxation during zazen meditation. By apply-ing EEG spectral analysis (Etevenon, Hen-rotte & Verdeaux, 1973), I discovered that,in the course of zazen meditation, there is ahighly hypovariable alpha peak correspondingto a sharp resonant alpha spectral peak to-gether with alpha amplitude modulation; thiscan easily be seen by the EEG envelope. In asubsequent publication (Ribemont, Etevenon& Giannella, 1979), I put forward a methodof ”zoom-FFT” for studying the alpha rangeand looking for hidden sharp frequency spec-tral EEG generators. This idea has recentlybeen taken up by Tognoli (Tognoli et al., 2007)via a new application of complex EEG Time-Frequency Analysis (complex Morlet wavelet),although it has not yet been applied to thestudy of EEGs of zazen meditators. I went onto pioneer the application of a Hilbert trans-form to EEG instead of a Fourier transformfor revealing the amplitude modulation of EEG(AM-EEG) (Etevenon, 1977); this new tech-nique was subsequently adopted in neuropsy-chology protocols (Etevenon, et al. 1999), al-though not yet in studies of Zen meditation.These innovative methods of EEG analysiscould now be applied to ongoing studies ofmeditation in the neurosciences.
Studies of Zen meditation have continuedsince then. James Austin (1999, 2006)published an essay on ”Zen and the Brain:Toward an Understanding of Meditation andConsciousness”, and this was followed in2006 by an important paper on ”Zen-Brainreflections”, linking modern neuroscience re-search findings with Eastern philosophy fromthe perspective of the unique Zen experi-ence. Subsequently, Kahana (2001, 2006)discussed the two rhythms that evidence par-
ticularly strong behavioral correlates: the 4–8Hz theta rhythm and the 30–80 Hz gammarhythm. It has been demonstrated that thetatrains occurring during zazen meditation maybe related to a working memory of maintaininga stable and steady zazen posture. Gammarhythm is perhaps simply masked by the highalpha rhythm which can also be looked uponas an inhibition of visual areas. During zazen,when attention is mostly directed inwards inorder to monitor body sensations and the flowof ongoing thoughts, alpha activity may alsobe enhanced, as Ray and Cole point out in thetitle of their paper (1985), because ”EEG al-pha activity reflects attentional demands, andbeta activity reflects emotional and cognitiveprocesses”. Lehmann and his collaborators inZürich (Lehmann et al, 2001; Tei et al., 2009)are also developing this line of research bymeans of new brain ”EEG source” imagingtechniques (LORETA), comparing meditatorswith non-meditators, and also altered states ofconsciousness with the experience of the self.
We need more research on this topic, becausemeditation studies may well give rise to key in-dicators that will enable the neurosciences toact as a bridge between different fields of ourcurrent knowledge.
Recent studies of meditation in theneurosciences
Francisco Varela (Varela, 1979, 1995, 1999;Varela & Shear, 1999; Varela, Thomson &Rosch, 1991) was a Chilean biologist, philoso-pher and neuroscientist who, together with hismentor Humberto Maturana, brought the con-cept of autopoiesis into biology (Varela & Mat-urana, 1980). He became a Tibetan Buddhist
2010, 3 (1) Kognitive Neurophysiologie des Menschen 11

P. Etevenon — ”Modified States of Consciousness” revisited
in the 1970s. In 1986, he settled in France,where he first taught cognitive science andepistemology at the CREA in the École Poly-technique, and neuroscience at the Universityof Paris. From 1988 until his death in 2001he led a research group at the LENA-CNRSwith Jacques Martinerie (Centre National deRecherche Scientifique; Varela et al., 2001).The LENA had been founded and headedby Antoine Rémond at the Pitié-Salpêtrièrehospital in Paris; Rémond pioneered studiesin quantitative EEG of spatio-temporal EEGmaps and evoked potentials (Remond, 1971-1978). In his 1995 keynote paper, Varela de-vised a new EEG method for analyzing brainwaves involving ”Resonant cell assemblies: Anew approach to cognitive functions and neu-ronal synchrony ”. He brought together suc-cessive time-events of 500 ms duration, whichLehmann later considered to be like ”quanta ofthoughts” of 100ms minimal epochs (Lehmannet al., 1998), brain electric microstates andmomentary-conscious mind states. Varelaassociated these time-events with behavioralevents, size of neuronal assemblies, EEG co-herence measurement in the high frequencygamma band, and topography of synchronybetween neuronal assemblies.
This EEG neural synchrony method was soonapplied in Paris to complex psychophysiolog-ical tasks (Lutz, 2002; Lutz et al., 2002), to-gether with a first-person approach to neu-rophenomenology. When Antoine Lutz wentto the United States, he continued this ap-proach through his studies of Buddhist medi-tators in Madison with Davidson (Brefczynski-Lewis, Lutz et al., 2007; Lutz et al., 2004,2007). In his 2004 PNAS paper, Lutz et al.developed the idea that ”long-term meditators
self-induce high-amplitude gamma synchronyduring mental practice”. I have reviewed thisarticle – which is highly technical – in the firstpart of ”States of consciousness” in my mostrecent work (Etevenon & Santerre, 2006), to-gether with 17 other major new papers onmeditation studies in the neurosciences, in-cluding that by Sara Lazar (Lazar et al., 2005).In her papers, Sara Lazar, working in Bostonin collaboration with Herbert Benson’s group,noted (2000) that in applying functional mag-netic resonance imaging (fMRI) ”the practice ofmeditation activates neural structures involvedin attention and control of the nervous sys-tem”. Later, in 2005, by applying magneticresonance imaging (MRI), they discovered thatthe ”meditation experience is associated withincreased cortical thickness” when ”meditationinsight” is being practiced; this implies that”cortical plasticity is associated with meditationpractice” (op. cit.).
Nowadays, as in the 1970s, meditation isagain being studied in the neurosciences, es-pecially in brain imagery laboratories. Re-cent findings open up new horizons either withrespect to our fundamental understanding ofwhat takes place or for practical purposes,such as alternative treatments in medicine andpsychotherapy, including maintaining the well-being of mind and body during the aging pro-cess. New theses are being published alongthese lines, while the history of meditation re-search (Cahn, 2007) still considers meditationto be an ”altered state of consciousness”. Ihave already argued that meditation has moreto do with ”voluntary controlled states of con-sciousness” (Henrotte, Etevenon & Verdeaux,1972; Etevenon, Henrotte & Verdeaux, 1973;Etevenon & Santerre, 2006), which itself is part
12 Human Cognitive Neurophysiology 2010, 3 (1)

P. Etevenon — ”Modified States of Consciousness” revisited
of a broader set of ”modified states of con-sciousness”. My principal argument is thatmeditators can enter into a meditative state,stay there if they like for some considerabletime, and also leave it whenever they wantto – whereas non-meditators will usually en-ter into the first stages of sleep after remain-ing quiet and sitting with their eyes closed for12 minutes. Cahn and Polich (2006) havepublished a paper on ”Meditation States andTraits: EEG, ERP, and Neuroimaging Stud-ies” and, more recently, one on the vipassanaBuddhist type of meditation (Cahn & Polich,2009, 2010; Cahn, Delorme & Polich, 2010),changing the P300 Event-Related Brain Po-tential. An additional paper on even morerecent findings on meditation and the neuro-sciences – from basic research to clinical prac-tice – is forthcoming (Braboszcz, Hahusseau& Delorme, 2010), and new articles and pa-pers by Davidson and Lutz will constantly up-date the state of the art and our developingscientific knowledge of meditation. New ma-terial is regularly being published on this topic,such as that by Andrew Newberg (Newberg,d’Aquili & Rause, 2002; Newberg & Waldman,2009) who coined the word ”neurotheology” af-ter studying the effect of praying on the EEG.Newberg’s research has been criticized fromtwo main perspectives. Firstly, from a religiouspoint of view, the study of practices such asmeditation cannot be extrapolated as such tothe broader area of religious and spiritual phe-nomena; secondly, from a non-religious per-spective, Newberg has been criticized for notin the end reducing religion to brain function-ing, a view held by some other neuroscien-tists. This debate has been going on sincethe 1970s, but the increasing amount of new
findings concerning the specificity of medita-tive states is making it more and more difficultto consider meditation as simply a pathologi-cal hallucinatory and internal behavioral expe-rience.
Conclusion and furtherperspectives
Results obtained over the previous ten years(Etevenon & Santerre, 2006) tend to indicatethat two categories of meditation are hence-forth to be taken into consideration.
Meditation ”with an object” involves sev-eral techniques: zazen, which begins withconcentration on respiration and posture; res-piratory yoga techniques, such as pranayama;repeated phrases like prayers or sanskritmantras (T.M.), in which auditory stimulationis maintained thanks to sustained atten-tion; visually-reinforced stimuli when focusingsteadily on a point, the light from a candle, areligious picture or icon, a mandala, a yantraor a specific inner visualization; and othermeditation techniques such as kundalini yoga(Silburn, 1983), yoga nidra, vipassana (Ri-card, 2008; Ricard & Thuan, 2000), ”insight”meditation, etc.
This kind of ”meditation with an object” ac-tivates the brain areas and networks relatedto the focused ”object” and is represented byactivated zones in 2D or 3D brain images byhigh resolution EEG with source localization(Brefczynski-Lewis, Lutz et al., 2007; Lutz etal., 2004), fMRI and MRI techniques (Lazar S.et al., 2000, 2005), or PET scan studies (ref-erences from the Institute of Noetic Sciences:IONS Bibliography ). These activated areascan be both cortical and sub-cortical.
2010, 3 (1) Kognitive Neurophysiologie des Menschen 13

P. Etevenon — ”Modified States of Consciousness” revisited
Meditation ”without an object” correspondsto techniques such as advanced zazen med-itation – hishiryo, amala, satori, sunyatastates of high consciousness as describedby Deshimaru (Deshimaru and Chauchard,1976); samadhi states and higher states ofconsciousness (Fischer, 1971, 1975, 1976;Gebser, 1949, 1985; Sri Aurobindo, 1970);Shiva consciousness in the Shivaïsm of Kash-mir (Silburn, 1970, 1977, 1981-1985, 1983);universal compassion in Buddhism (Silburn,1977; Ricard, 2008); and so forth.As a counterpart to the preceding type, ”med-itation without an object” relaxes and deacti-vates the brain areas and networks related toforgotten or unfocused objects such as the ‘I’,as well as brain areas linked to time and/orspace percepts and concepts. This is repre-sented by deactivated zones in 2D or 3D brainimages together with increased synchrony be-tween large assemblies and populations ofneurons ((Brefczynski-Lewis, Lutz et al., 2007;Lutz et al., 2004). These deactivated areascan be both cortical and sub-cortical.
Further perspectives
A whole new philosophical vocabulary hascome to the fore: neurophenomenology,neurophilosophy, neurotheology and, morerecently, neuroeconomy, neurocybernetics,neuroethics, neuroesthetics, etc. Each ofthese terms argues in favor of a particularconception of states of consciousness, basedupon and explained by specific relationshipsand connectivity in the neurodynamics of thebrain and its networks.Walter Freeman, who set up the Freeman Lab-oratory for Nonlinear Neurodynamics in Berke-ley, is very much in the forefront of these theo-
retical ventures; he has written some highly im-portant seminal books (Freeman, 1975, 1995,2000), together with a significant number of re-search and teaching articles.
Matters of free will and determinism have beenunder discussion for some time now (Fischer,1967, 1971, 1975, 1976). Lazar (Lazar etal., 2000, 2005) formulated one such ques-tion thus: is the individual brain predisposedto being particularly conducive to meditationor is it the practice of ”insight meditation” itselfwhich increases the thickness of specific brainareas? That, of course, evokes the well-knownquestion about which came first, the chickenor the egg; in 1965 at the MIT in Boston, Iwas told that this was the kind of question thatonly a bird could ask! Multiple causality isquite possible; in my D.Sc. thesis (1977), I ap-plied the sophisticated but linear Gersch anal-ysis (Gersch, 1972) of ”causality and driving inelectrophysiological signal analysis” and wenton to put forward some hypotheses on parallelversus serial functioning of large assemblies ofneurons (Etevenon, 1992).
New protocols of research into meditativestates may be developed and studied, bring-ing together data from the latest techniques ofbrain imagery in order to enhance our under-standing of ”modified states of consciousness”(M.S.C.), including not only meditation butalso shamanic trance (cf. Pierre Flor-Henry’sinnovative recordings of Corine Sombrun inEdmonton [personal communication]), hyp-nosis (Rainville and Price, 2003; Grant &Rainville, 2005), N.D.E. experiences, neuro-feedback studies, ”lucid dreaming” as a formof ”proto-consciousness” (Bouchet, 1994;LaBerge, 2000), REM states in sleep anddreaming (Hobson, 2009), etc.
14 Human Cognitive Neurophysiology 2010, 3 (1)

P. Etevenon — ”Modified States of Consciousness” revisited
In addition, philosophical works have beenpublished by Sri Aurobindo (1949, 1970), byJean Gebser (1949, 1985) and by those whoinitiated the present trend towards integralistphilosophy (Combs, 2002; Feuerstein, 1987).In their reflections on the stages of evolutionof human beings, they argue that after ourpresent ”mental structure (Jean Gebser) orplane (Sri Aurobindo) of consciousness” an in-tegral structure or supermind plane of con-sciousness will gradually evolve. We maytherefore look forward to further discoveriesarising from prospective studies of meditativestates by the neurosciences; these may wellindicate a positive evolution of human beingsand society, leading towards the developmentof integral or supermind consciousness. AsFrançois Rabelais put it, ”science sans con-science n’est que ruine de l’âme” (”Knowledgewithout conscience is but the ruin of the soul”).
Acknowledgements
This review is based on a plenary lecture, en-titled ”From states of meditation studied in theneurosciences to voluntary controlled states ofconsciousness”, that I was invited to give inApril 2009 at the ”Deuxième Colloque interna-tional transdisciplinaire sur les Hallucinationset autres États Modifiés de Conscience dans laPhilosophie et les Sciences Cognitives” (ParisSecond Spring Symposium on Hallucinationsin Philosophy and Cognitive Science), orga-nized by G. Dumas, J. Gonzalez, A. Lehmannand J. Kevin O’Reagan, whom I wish to thankfor their invitation.I would like also to express my gratitude toAlexandre Lehmann for his support, construc-tive comments and corrections, to Nicolas dela Boutresse for his helpful criticism and obser-
vations on an earlier version of this paper, andto David Alcorn for his linguistic and semanticrevision of this final version.Last but not least, the many encouraging com-ments by Wolfgang Skrandies, the editor ofthis scientific journal, well known for its open-mindedness and freedom of expression, andthe positive input of the three reviewers, madeit possible for me to finalize this paper for pub-lication.
www.sgdl-auteurs.org/pierre-etevenon/
2010, 3 (1) Kognitive Neurophysiologie des Menschen 15

P. Etevenon — ”Modified States of Consciousness” revisited
References
Anand B.W., Chhina G.S. & Singh B. (1961)Some Aspects of ElectroencephalographicStudies in Yogis. Electroenceph. Clin. Neuro-physiol, 13, 452-456.
Anceau F. (1999) Vers une étude objective dela conscience. Hermès Science Publications,Paris.
Austin J.H (1999) Zen and the Brain: Toward anUnderstanding of Meditation and Conscious-ness. MIT Press Books, Cambridge.
Austin J.H. (2006) Zen-Brain reflections. MITPress Books, Cambridge.
Bagchi B.K. & Wenger M.A. (1957) Electrophys-iological Correlates of Some Yogi Exercises.Electroenceph. Clin. Neurophysiol, 7, 132-149.
Banquet J.P. (1973) Spectral analysis of theEEG in meditation. Electroenceph. Clin. Neu-rophysiol, 35, 143-151.
Beaubernard C. (2009) Rêves Récurrents. Voieroyale pour l’étude et l’explication des rêves. Àpropos d’un enfant. Préface de Michel Jouvet.Sciences, Neurophysiologie, Éditions Publi-Book Université, Paris.
Benson H. (1975) The Relaxation Response.William Morrow and Co.; (1976, 2000) AvonBook, New York.
Benson H. (1987) Your Maximum Mind. TimesBooks/Random House, New York.
Benson H. (1997) The relaxation response: ther-apeutic effect, Science, 278, 1694-1695.
Benson, H., Lehmann, J.W., Malhotra, M.S.,Goldman RF, Hopkins J & Epstein MD. (1982)Body Temperature Changes during the Prac-tice of g Tum-mo (Heat) Yoga, Nature, 295,234-236.
Benson H. & Proctor W. (1985) Beyond the
Relaxation Response, Berkley Books, Pen-guin Putnam, New York; in French (2008) Leprincipe de l’étincelle, Sand-Tchou, Paris.
Benson H & Proctor W. (2003) The breakoutprinciple. Scribner, New York.
Benson H. & Stark M. (1996) Timeless Healing:The Power and Biology of Belief. Scribner,New York.
Bouchet C. Le rêve lucide. (1994) Thèse deDoctorat d’État ès Lettres, Paris Sorbonne.http://www.svabhinava.org/friends/ChristianBouchet
/TOC-English-frame.php
Braboszcz C., Hahusseau S. & Delorme A.(2010) Meditation and neuroscience : frombasic research to clinical practice. In CarlstedR. (Ed.): Integrative Clinical Psychology, Psy-chiatry and Behavioral Medicine Perspectives.Practices and Research., Springer, Berlin.
Brefczynski-Lewis J.A., Lutz A., Schaefer H.S.,Levinson D.B. & Davidson R.J. (2007) Neuralcorrelates of attentional expertise in long-termméditation practitioners. Proc. Natl. Acad.Sci., (PNAS), 104, 11483-11488.
Cahn B.R. (2007) Neurophysiologic Corre-lates to Sensory and Cognitive Processingin Altered States of Consciousness. Neuro-Science, San Diego (UCSD, PhD thesis).
Cahn B.R. & Polich J. (2006) Meditation Statesand Traits: EEG, ERP, and NeuroimagingStudies. Psychological Bulletin, 132, 180-211.
Cahn B.R. & Polich J. (2009) Meditation (Vipas-sana) and the P3a event-related brain poten-tial, International Bulletin of Psychophysiol-ogy, 72, 51-60.
Cahn B.R., Delorme A. Polich J. (2010). Occip-ital gamma activation during Vipassana medi-tation. Cognitive processing 11, 39-56.
Changeux J.P. (1983) L’homme neuronal. Fa-yard, Paris.
16 Human Cognitive Neurophysiology 2010, 3 (1)

P. Etevenon — ”Modified States of Consciousness” revisited
Combs A. (2002) The Radiance of Being: Un-derstanding the Grand Integral Vision. Liv-ing the Integral Life. Omega Book, New York,Paragon House, St Paul.
Coupey P. (2006) ZEN Simply Sitting a Zenmonk’s commentary on the Fukanzazengi ∼Universal Guide to the Practice of Zazen ∼ byMaster dogen (1200 - 1253). HOHM PRESS,Prescott, Arizona.
Coxeter H.S.M. (1974) Projective Geometry,second edition, Springer, New York.
Crick F. (1994) The Astonishing Hypothesis. TheScientific Search for the Soul. MacMillan Pub-lishing Company; French translation (1994):L’hypothèse stupéfiante, à la recherche scien-tifique de l’âme, Plon, Paris.
Dang-Vu T.T., Schabus M., Desseilles M., Al-bouy G., Boly M., Darsaud A., Gais S.,Rauchs G., Sterpenich V., Vandewalle G.,Carrier J., Moonen G., Balteau E., DegueldreC., Luxen A., Phillips C. & Maquet P. (2008).Spontaneous neural activity during humanslow wave sleep. Proc. Natl. Acad. Sci. 105,15160-15165.
Das, N. & Gastaut H. (1957). Variations dansl’activité électrique du cerveau, du cœur etdes muscles squelettiques au cours de laméditation et de l’extase yogique. [Variationsof the electrical activity of the brain, heart, andskeletal muscles during yogic meditation andtrance.] Electroenceph. clin. Neurophysiol.,Suppl. 6, 211–219.
Davidson J.M. (1976) The Physiology of Medi-tation and Mystical States of Consciousness.Perspectives in Biology and Medicine 19, 3,345-380.
Davidson J.M. & Davidson R.J. (Eds.) (1980)The Psychobiology of Consciousness.Plenum Press, New York.
Davidson R.J. & Harrington A. (Eds.) (2002).Consciousness. Harper and Row, New York.
Dehaene S., Changeux J.P., Naccache L. &Sackur J. (2006) Conscious, preconscious,and subliminal processing: a testable taxon-omy. Trends in Cognitive Sciences, 10, 204–211.
Deniker P. , Boissier J.R., Etevenon P., GinestetD., Peron-Magnan P. & Verdeaux G. (1974)Étude de pharmacologie clinique du Delta-9tétra-hydro-cannabinol chez des sujets volon-taires sains avec contrôle polygraphique.Thérapie, 29, 185-200, Paris.
Dennett D.C. (1991) Consciousness Explained.Little, Brown and Company; French Transla-tion (1993): La conscience expliquée, ÈditionsOdile Jacob, Paris.
Descamps M.A. (2006) Rencontres avec douzefemmes remarquables. Alphée, Monaco.
Deshimaru Taisen Maître. (1981) La pratique duzen, Spiritualités vivantes, Albin Michel, Paris.
Deshimaru Roshi Taisen & Chauchard P. (1976)Zen et cerveau, Le Courrier du livre, Paris.
Eccles J.C. (1989) Evolution of the Brain. Cre-ation of the Self. Routledge; French transla-tion (1992): Évolution du cerveau et créationde la conscience, Fayard, Paris.
Edelman G.M. (1992) Bright Air, Brillant Fire: Onthe Matter of Mind. Annen Lane, The PenguinPress. (French translation: Biologie de la con-science, Éditions Odile Jacob, Paris).
Eliade M. (1958) Yoga: Immortality and Free-dom. Translation by Trask W.R., Routledgeand Kegan, London.
Eliade M. (1967) From Primitives to zen. Harperand Row, New York.
Engelberg E. (1972) The Unknown distance.Harvard University Press, Cambridge, Mass.
Erigena John Scotus. Article by Dermot Moran
2010, 3 (1) Kognitive Neurophysiologie des Menschen 17

P. Etevenon — ”Modified States of Consciousness” revisited
in the Stanford Encyclopedia of Philosophy, inhttp://plato.stanford.edu/entries/scottus-eriugena/
Etevenon P. (1967) Effets de la S.L.I. sur lesaires cortico-visuelles du lapin. Latéralisationspécifique des réponses électroencéphalo-graphiques et variations aprés administra-tion d’amphétamine et pentobarbital. Noteprésentée par A. Fessard, C.R., C.R. Acad.Sci. , Paris, 265, 885-888.
Etevenon P. (1967) Effets de la stimulation en lu-mière intermittente (SLI) différentielle sur lesstructures visuelles du lapin. Variations del’électrogénèse et de la dominance cérébralesous l’action du LSD-25. Revue de MédecineAéronautique, 21, 35-47, Paris.
Etevenon P. (1974) Étude comparée des étatsde conscience dans la tradition Indienne. 55,17-39, n˚spécial, La Vie Médicale, Paris.
Etevenon P. (1974) Étude psychophysiologiquecomparée des états de sommeil et de médi-tation. 55, 7-14, n˚ spécial, La Vie Médicale,Paris.
Etevenon P. (1974) From modern science toSri Aurobindo’s integral knowledge, 206-211in K.R.S. Iyengar (Ed.), Sri Aurobindo. Acentenary tribute, International Seminar inNew Delhi, Sri Aurobindo Ashram Press,Pondicherry.
Etevenon P. (1977) Étude méthodologiquede l’électroencéphalographie quantita-tive. Application à quelques exemples.Thèse de doctorat d’état ès sciencesnaturelles, soutenue le 29.11.1977 àl’Université Pierre et Marie Curie Paris VI,303 p., Copédith, Paris, http://www.faustine-
g.com/Romain/These_P_Etevenon.pdf
Etevenon P. (1978) Effects of cannabis on hu-man EEG, Adv. Biosci., 22-23, 659-663.
Etevenon P. (1984) Les Aveugles éblouis. Les
états limites de la conscience. Albin-Michel,Paris.
Etevenon P. (1985) Électroencéphalogra-phie quantitative, Encyclopédie Médico-chirurgicale, 3717 dans Psychiatrie, 1èreed. , A10, 4, II.
Etevenon P. (1986) Applications and perspec-tives of EEG cartography [mental calculationarithmetic task in a group of normal subjects].Chapter 6, 115-142 in F. H. Duffy Ed., Topo-graphic Mapping of Brain Electrical Activity,Butterworths Pub., Stoneham, Mass.
Etevenon P. (1986) La caverne de Platon. Oucartographie d’une nuit de sommeil et derêve. Movie on videocassette VHS-PAL-SECAM, 25 minutes, N˚ CNC edv 169,CNRS DIFFUSION, Meudon, internet sites :http://videotheque.cnrs.fr or http://www.bium.univ-
paris5.fr/medecine/video.pdf
Etevenon P. (1987) Du rêve à l’éveil. Basesphysiologiques du sommeil, Albin-Michel,Paris.
Etevenon P. (1989) EEG dynamic cartographyof wakefulness sleep and dreams : A movie.329-334 in K. Maurer (Ed.) TopographicBrain Mapping of EEG and Evoked Potentials,Springer, Berlin.
Etevenon P. (1989) Dynamic EEG mappingof vigilance and dream processes. Hobsonand Mc Carley Hypothesis revisited, 73-75 inHorne J. (Ed.) “Application of brain ImagingMethods: Sleep ’88“, G. Fischer, Stuttgart.
Etevenon P. (1990) L’Homme Éveillé. Paradoxesdu sommeil et du rêve. Sand-Tchou, Paris.
Etevenon P. (1992) Hypercube model (gamma-4) and embedded regular hypercubes ofhigher dimensions applied in psychology andneurosciences. Actes du 13ème CongrèsInternational de Cybernétique, Association
18 Human Cognitive Neurophysiology 2010, 3 (1)

P. Etevenon — ”Modified States of Consciousness” revisited
Internationale de Cybernétique, 716-720,Namur.
Etevenon P. (1993) Analyse quadri-dimensionnelle de l’électroencéphalogramme.Première application à un épisode de sommeilparadoxal chez l’Homme. Note présentée parM. Jouvet. C. R. Acad. Sci. Paris, 316, SérieIII, neurophysiologie, 267-274.
Etevenon P. (1994) Éveil, attention, concen-tration. . . transformation, Revue Française deyoga, 9, Dhâranâ, 91-103, Paris.
Etevenon P. (1999) Les états de consciencedans la tradition indienne, Les Carnets duyoga, 197, 29-32, U.N.Y., Paris.
Etevenon P. (1999) Yoga et relaxation, Les Car-nets du yoga, 201, 10-15, U.N.Y., Paris.
Etevenon P. (2007) Un scientifique épris deyoga, Infos-Yoga, 61, 16-17, et 62, 39, Lalleu,35320, France. http://auriol.free.fr/yogathera/
transpersonnel/Article_Infos-Yoga_PE.htm
Etevenon P. (2007) Les neurosciences etle transpersonnel, Bulletin du Transper-sonnel, N˚ 86, 3-16, AssociationFrançaise du Transpersonnel (A.F.T.),Paris. http://auriol.free.fr/yogathera/ transper-
sonnel/Neurosciences_PE.htm
Etevenon P. & Boissier J.R. (1972) LSD ef-fects on signal-to-noise ratio and lateralisationof visual cortex and lateral geniculate duringphotic stimulation. Experientia, 28, 11, 1338 –1340.
Etevenon P. & Boissier J.R. (1974) Approchesthéoriques de la schizophrénie. La Revue deMédecine, 15, 643-651.
Etevenon P., Gaches J., Debouzy C., GueguenB. & Peron-Magnan P. (1985) EEG cartogra-phy I. By means of mini or microcomputers.Reliability and interest of this electrical non-invasive brain imagery. Neuropsychobiology,
13, 69 - 73.
Etevenon P. & Guillou S. (1986) EEG cartogra-phy of a night of sleep and dreams, Neuropsy-chobiology, 16, 146-151.
Etevenon P., Henrotte J.G. & Verdeaux G. (1973)Approche méthodologique des états de con-science modifiés volontairement (Analysespectrale statistique). Rev. EEG Neurophys-iol. clin., Paris, 3, 232–237.
Etevenon P., Lebrun N., Clochon P., Perchey G.,Eustache F. & Baron J.-C. (1999) High tem-poral resolution dynamic mapping of instanta-neous EEG amplitude modulation after tone-burst auditory stimulation, Brain Topography,12, 129-137.
Etevenon P., Pidoux B. & Rioux P. (1979) Strat-egy of statistical spectral analysis in drug stud-ies. A methodological synopsis, 383-391 inSaletu B. (Ed.) Neuropsychopharmacology,Pergamon Press, Oxford.
Etevenon P., Pidoux B., Rioux P., Peron-MagnanP., Verdeaux G. & Deniker P. (1979) Intra- andInterhemispheric EEG differences quantifiedby spectral analysis. Comparative study oftwo groups of schizophrenics and a controlgroup. Acta Psychiatrica Scandinavica, 60,57-68. Published Online: 23 Aug 2007, DOI
10.1111/j.1600-0447.1979.tb00265.x, Wiley Inter-
science.
Etevenon P. & Santerre B. (2006) États de con-science Sophrologie et Yoga, Sand-Tchou,Paris. www.editions-tchou.com.
Etevenon P., Tortrat D., Guillou S. & WendlingB. (1985) Cartographie EEG au cours d’unetâche visuo-spatiale. Cartes moyennes etstatistiques de groupes. Rev. EEG. Neuro-physiol. Clin., 15, 139 - 147.
Fenwick, P., Donaldson S., Gillis L., Bushman,J., Fenton, G. W., Perry, I., Tilsley, C. & Ser-
2010, 3 (1) Kognitive Neurophysiologie des Menschen 19

P. Etevenon — ”Modified States of Consciousness” revisited
afinowicz, H. (1977) Metabolic and EEGchanges during transcendental meditation:An explanation. Biological Psychology 5,101-118.
Feuerstein G. (1987) Structures of Conscious-ness. The Genius of Jean Gebser. IntegralPublishing, Santa Rosa.
Fischer R. (Ed.) (1967) Interdisciplinary Per-spectives of Time. Annals N.Y. Acad.Sci.,138, 367-915.
Fischer R. (1971) A cartography of the ecstaticand meditative states. Science, 174 , 897-904.
Fischer R. (1972) On the arousal state-dependent recall of ’subconscious’ experi-ence stateboundness. The British Journal ofPsychiatry, 120, 159-172.
Fischer R. (1975) Transformations of con-sciousness. A cartography I. The perception-hallucination continuum. Confin. Psychiat.18, 221-244.
Fischer R. (1976) Transformations of conscious-ness. A Cartography II. The perception-hallucination continuum. Confin. Psychiat.19, 1-23.
Flor-Henry P. (2007) Personal Communicationand unpublished Lecture on ”Neurophysiolog-ical changes occurring during trance (Mon-golian induction): relationship to depersonal-ization, out-of-body experience, dissociation,and auto-hypnosis”. This was following first EEG
recordings on December 19, 2007 together with
spectral and LORETA analyses of the EEGs of
Corine Sombrun who is a French musician, writer
and a recognized shaman in Mongolia after 8 years
of training.
Freeman W. (1975) Mass Action in the NervousSystem. Academic Press, San Diego.
Freeman W. (1995) Societies of Brains.
Lawrence Erlbaum.
Freeman W. (2000) Neurodynamics. Springer,Berlin and http://sulcus.berkeley.edu/.
Freeman W. (2000) Reclaiming Cognition. Im-print Academic.
Gebser J. The Ever-Present Origin. (1985,1991) Part I, Part II, Ohio University Press,Athens; English translation of (1949, 1953)Ursprung und Gegenwart, I, II, DeutscheVerlags-Anstalt GmbH, Stuttgart.
Gellhorn E. & Kiely W.F. (1972) Mystical statesof consciousness: Neurophysiological andclinical aspects. The Journal of Nervous andMental Disease, 154, 399-405.
Gersch W. (1972) Causality or driving in elec-trophysiological signal analysis. Math. Bio-sciences. 14, 177-196.
Goldstein L. (1979) Is a man, a man, a man?(or: is an EEG, an EEG, an EEG?) Some re-marks on the homogeneity of normal subjects.Pharmakopsychiat. Neuropsychopharmakol.,12, 74-78.
Goldstein L. (1983) Brain function and behavior:on the origin and evolution of their relatioships.Adv. Biol. Psychiatry, 13, 75-79.
Goldstein L. (1983) Some EEG correlates ofbehavioral traits and states in humans. Re-search Communications in Psychology Psy-chiatry and Behavior, 8, 115-141.
Grant J.A. & Rainville P. (2005) Hypnosis andmeditation: similar experiential changes andshared brain mechanisms.Med Hypotheses,65, 625-626.
Green E.E., Green A.M. & Walters E.D. (1970)Voluntary control of internal states: Psycho-logical and physiological, Journal of Transper-sonal Psychology, 1-26.
Günther G. (1967) Time, Timeless Logic andSelf-Referential Systems,in Interdisciplinary
20 Human Cognitive Neurophysiology 2010, 3 (1)

P. Etevenon — ”Modified States of Consciousness” revisited
Perspectives of Time, R. Fischer (Ed.), Ann.N.Y. Acad. Sc., 138, 396-406.
Günther G. & von Foerster H. (1967) The logicalstructure of evolution and emanation, in Inter-disciplinary Perspectives of Time, R. Fischer(Ed.), Ann. N.Y. Acad. Sc., 138, 874-891.
Hebert R. & Lehmann D. (1977) Theta bursts:an EEG pattern in normal subjects practic-ing the transcendental meditation technique.Electroenceph. Clin. Neurophysiol. 42, 397-405.
Henrotte J.G. (1969) Yoga et biologie, Atomes,265, 283-292.
Henrotte J.G. (2001) Entre Dieu et Hasard. Unscientifique en quête de l’Esprit, L’Harmattan,Sciences et Société, Paris.
Henrotte J.G., Etevenon P. & Verdeaux G. (1972)Les états de conscience modifiés volontaire-ment. 3, 1100-1102, La Recherche, Paris.
Hirai T. (1960) Electroencephalographic studyon the Zen meditation (Jap.) Psychiat. Neurol.Jap., 62, 76-105.
Hirai T. (1974) The Psychophysiology of Zen.Igaku Shoin, Tokyo.
Hobson A.J. (2009) REM sleep and dreaming:towards a theory of protoconsciousness. Na-ture Reviews Neuroscience, 10, 803-813.
Hoffman J.W., Benson H., Arns, P.A., StainbrookG.L., Landsberg G.L., Young, J.B. & Gill, A.(1982) Reduced sympathetic nervous systemresponsivity associated with the relaxation re-sponse, Science, 215, 190-192.
”Institute of Noetic Sciences” : IONS Bibliogra-phy on Meditation, http://biblio.noetic.org/
James W. (1902) The Varieties of Religious Ex-perience. Longmans, New York, reprinted1916, Longmans Green and 1958, New Amer-ican Library, New York.
Jouvet M. (2009) Symposium International
SFRS/WASM pour l’année du cinquantièmeanniversaire de la découverte du sommeilparadoxal, 7-10 janvier 2009, Lyon.
Kahana M.J. (2001) Theta returns. CurrentOpinion in Neurobiology, 11, 739-744.
Kahana M.J. (2006) The cognitive correlates ofhuman brain oscillations, The Journal of Neu-roscience, 26, 1669-1672.
Kasamatsu A. & Hirai T. (1963) Science ofzazen. Psychologia, 6, 86-91.
Kasamatsu A. & Hirai T. (1966) An electroen-cephalographic study on the Zen meditation(Zazen), Folia Psych. Neurol. Japon, 20, 315-336.
Kazamatsu A., Okuma T. & Takenaka S. (1957)The EEG of ‘Zen’ and ‘Yoga’ practitioners.Electroenceph. Clin. Neurophysiol., Suppl. 9,51-52.
Kazamatsu A., Okuma T., Takenaka S., KogaE., Ikeda K. & Sugiyama H. (1958) The EEGof Zen and Yoga practitioners, Electroenceph.Clin. Neurophysiol., 10, 193, (abstract).
” La conscience, Exploration au centre ducerveau ”, (2008) Les Dossiers de LaRecherche, L. Naccache (Ed.), N˚30, Paris.La conscience was also the main topic of a re-
cent scientific meeting where ” Consciences et
Neuropsychologie ” (2010) was the first thema of
the ” Cinquièmes Rencontres de Neurologie Com-
portementale ” organised 4th February 2010 in
Paris, with S. Dehaene chairman and L. Naccache,
S. Dehaene and M. Jeannerod successive main
speakers.
LaBerge, S. (2000). Lucid dreaming: Evidenceand methodology. Behavioral and Brain Sci-ences 23, 962-963. www.lucidity.com
Lapassade G. (1987) Les États modifiés de laconscience, PUF, Paris.
Lazar SW, Bush G, Gollub RL, Fricchione GL,
2010, 3 (1) Kognitive Neurophysiologie des Menschen 21

P. Etevenon — ”Modified States of Consciousness” revisited
Khalsa G & Benson H. (2000) Functional brainmapping of the relaxation response and med-itation. NeuroReport, 11, 1581-1585.
Lazar S.W., Kerr C.E., Wassermann R.H., GrayJ.R., Greve D.N., Treadway M.T., McGarveyM., Quinn B.T., Dusek J.A., Benson H., RauchS.L., Moore C.L. & Fischl B. (2005) Meditationexperience is associated with increased corti-cal thickness, NeuroReport,16, 1893-1897.
Lebrun N., Clochon P., Etevenon P., EustacheF. & Baron J.C. (1998) Effect of environmen-tal sound familiarity on dynamic neural acti-vation/inhibition patterns: An ERD mappingstudy. NeuroImage, 8, 79-92.
Lebrun N., Clochon P., Etevenon P., Lambert J.,Baron J.C. & Eustache F. (2001) An ERD map-ping study of the neurocognitive processes in-volved in the perceptual and semantic analy-sis of environmental sounds and words. Cog-nitive Brain Research, 11, 235-248.
Lehmann D., Faber P.L., Achermann, P., Jean-monod, D., Gianotti, L.R.R. & Pizzagalli, D.(2001) Brain sources of EEG gamma fre-quency during volitionally meditation-induced,altered states of consciousness, and experi-ence of the self. Psychiatry Res.: Neuroimag-ing 108, 111-121.
Lehmann D., Strik W.K., Henggeler B., KoenigT. & Koukkou M. (1998) Brain electric mi-crostates and momentary conscious mindstates as building blocks of spontaneousthinking: I. Visual imagery and abstractthoughts, Int. J. Psychophysiol. 29, 1-11.
Lutz A. (2002) Pour une Approche Neu-rophénoménologique des Bases Neuro-dynamiques de l’Expérience Consciente:Application à un protocole de Vision Stéréo-scopique. Thèse de l’Université Paris VI, 118p.
Lutz A., Dunne J.D. & Davidson R.J. (2007),Meditation and the Neuroscience of Con-sciousness : An introduction, in The Cam-bridge Handbook of Consciousness, chap.19, 497-549.
Lutz A., Lachaux J.P., Martinerie J. & Varela F.J.(2002) Guiding the study of brain dynamics byusing first-person data: Synchrony patternscorrelate with ongoing conscious states dur-ing a simple visual task. Proc. Natl. Acad.Sci. USA, (PNAS), 99, 1586-1591.
Lutz A., Greischar L.L., Rawlings N.B., Ricard M.& Davidson R.J. (2004) Long-Term meditatorsself-induce high-amplitude gamma synchronyduring mental practice, Proc. Natl. Acad. Sci.USA, (PNAS), 101, 16369-16373.
Lutz A., Slagter H.A., Dunne J.D. & DavidsonR.J. (2008) Attention regulation and monitor-ing in méditation, Trends in Cognitive Science,12, 163-169.
Mahowald M.W. & Schenck C.H. (1992) Dissoci-ated States of Wakefulness and Sleep. Neu-rology, 42, Suppl 6, 44-51.
Maslow A. (1970) Religion, Values and Peak Ex-periences. Penguin Books, London.
Masui J. (1952) De la vie intérieure, Documentsspirituels, Cahiers du Sud, Paris.
Metzinger, T. (1995) Bewußtsein Beiträge ausder Gegenwartsphilosophie. Paderborn:mentis; Conscious Experience. Thorver-ton, UK: Imprint Academic & Paderborn:mentis; available as http://www.philosophie.uni-
mainz.de/metzinger/
Mind and Life I (1995) Passerelles, entretiensavec des scientifiques sur la nature de l’esprit,Poche, Albin Michel, Paris.
Mind and Life II (1989) Le Pouvoir de l’esprit,Entretiens avec des scientifiques, Dalaï LamaXIV; (2000) Editions Fayard et Pocket, Paris.
22 Human Cognitive Neurophysiology 2010, 3 (1)

P. Etevenon — ”Modified States of Consciousness” revisited
Mind and Life III (1990) Quand l’esprit dialogueavec le corps, Daniel Goleman (Ed.); 1998Guy Trédaniel (Ed.), Paris.
Mind and Life IV (1992) Francisco J. Varela,Claude B. Levenson, Dormir, rêver, mourir,explorer la conscience avec le Dalaï Lama;(1998) Nil éditions, Paris.
Mind and Life VIII (2000) : Dalaï-Lama, DanielGoleman, Surmonter les émotions destructri-ces, Un dialogue scientifique avec le DalaïLama ; (2003) Editeur Robert Laffont, 2003& Pocket, Paris.
Monod J. (1973) Le hasard et la nécessité. PointSeuil, Paris
Murphy M. (1992) The Future of the Body. J.P.Tarcher/Putnam, New York.
Murphy M. & Donovan S. (1983) A bibliogra-phy of meditation theory and research: 1931-1983, Journal of Transpersonal Psychology,15, 181-228.
Murphy M. & Donovan S. (1999-2009) ThePhysical and Psychological Effects of Medi-tation. A Review of Contempory Research.Updated by E. Taylor as http://www.noetic.org/
research/ medbiblio/ch_intro1.htm.
Naccache L. (2006) Le nouvel inconscient:Freud, le Christophe Colomb des neuro-sciences. Éditions Odile Jacob, Paris.
Naranjo C. & Ornstein N.E. (1971) On the psy-chology of meditation. Viking press, NewYork.
Newberg A., d’Aquili E. & Rause V. (2002) WhyGod Won’t Go Away: Brain Science and theBiology of Belief, Ballantine Book, RandomHouse, New York; (2003) in French : Dieu nedisparaîtra pas. Quand la science explique lareligion, Sully.
Newberg A. & Waldman, M.R. (2009) How GodChanges Your Brain: Breakthrough Findings
from a Leading Neuroscientist, BallantineBook, Random House, New York.
Orme-Johnson D.W. & Haynes C.T. (1981)EEG phase coherence, pure consciousness,creativity, and TM-sidhi experiences, Interna-tional J. of Neuroscience, 13, 211-217.
Ornstein R.E. (1972) The psychology of con-sciousness. W.H. Freeman, San Francisco.
Penrose R. (1994) Shadows of the Mind. OxfordUniversity Press; French translation (1995):Les ombres de l’esprit, InterEditions.
Pfurtscheller G. & Aranibar A. (1977) Event-related desynchronization detected by powermeasurements of scalp EEG. Electroenceph.Clin. Neurophysiol. 42, 817-826.
Pfurtscheller G. & Lopes da Silva F.H. (1999)Event-related EEG/EMG synchronization anddesynchronization : basic principles. Clin.Neurophysiol. 110, 1842-1857.
Rainville P. & Price DD. (2003) Hypnosis phe-nomenology and the neurobiology of con-sciousness. Int J Clin Exp Hypn., 51, 105-129.
Ray W.J. & Cole H.W. (1985) EEG alpha activityreflects attentional demands, andbBeta activ-ity reflects emotional and cognitive processes.Science, 228, 750-752.
Remond A. (1971-1978) Handbook of Elec-troencephalography and Clinical Neurophys-iology. Elsevier Scientic Pub. Co., Amster-dam.
Ribemont B., Etévenon P. & GiannellaF (1979) Une méthode d’approche enélectroencéphalographie : l’analyse deFourier en bande étroite. Septième Col-loque sur le traitement du signal et sesapplications GRETSI, Nice, 101-104.http://documents.irevues.inist.fr/ bitstream/handle/
2042/10289/AR101.pdf?sequence=1
Ricard M. (2008) L’art de la méditation, Nil édi-
2010, 3 (1) Kognitive Neurophysiologie des Menschen 23

P. Etevenon — ”Modified States of Consciousness” revisited
tions, Paris.
Ricard M. & Thuan T.X. (2000) L’infini dans lapaume de la main. Du Big Bang à l’Éveil, NilÉditions / Fayard, Paris.
Roberts S. (2006) King of Infinite Space. Don-ald Coxeter, the Man Who Saved Geometry,Walker & Co., New York.
Silburn L. (1970) Abhinavagupta. Publication In-stitut de Civilisation Indienne, Paris.
Silburn L. (Ed.) (1977) Le Bouddhisme. Fayard,Paris.
Silburn L. (Directeur de collection) (1981-1985)Revue Hermès. 4 tomes, Les deux Océans,Paris.
Silburn L. (Ed.) (1983) La Kundalini. Les deuxOcéans, Paris.
Sri Aurobindo. (1970) Birth Centenary Li-brary, 30 volumes, Sri Aurobindo ashram,Pondicherry, All India Press, India.
Sri Aurobindo, (1949, 1955, 1977) The Life Di-vine (in French La Vie Divine, 2005), Sri Au-robindo Ashram Press, Pondicherry, India.
Synge J.L. (1951) Science: Sense and Non-sense, Jonathan Cape, London.
Tart C.T. (1969) Altered States of Conscious-ness. John Wiley and Sons, New York.
Tei, S., Faber, P.L., Lehmann, D., Tsujiuchi, T.,Kumano, H., Pascual-Marqui, R.D., Gianotti,L.R.R. & Kochi, K. (2009) Meditators and non-meditators: EEG source imaging during rest-ing. Brain Topography. 22, 158-165.
Timmons B. & Kamiya J. (1970) The psychol-ogy and physiology of méditation and relatedphenomenon : a bibliography, Journal ofTranspersonal Psychology, 2, 41-59, 1970.
Timmons B. &Kanellakos D. (1974) The Psy-chology and Physiology of Meditation and Re-lated Phenomena: Bibliography II. Journal ofTranspersonal Psychology, 6, 32-38.
Tognoli E., Lagarde J., DeGuzman G.C. & KelsoJ.A.S. (2007) The phi complex as a neuro-marker of human social coordination, Proc.Natl. Acad. Sci. USA, (PNAS), 104, 8190-8195.
Varela F. (1979) Principles of Biological auton-omy, North-Holland.
Varela F.J. (1995) Resonant cell assemblies : Anew approach to cognitive functions and neu-ronal synchrony, Biol. Res., 28, 81-95.
Varela F.J. (1999) Dormir, rêver, mourir. Explorerla conscience avec le Dalaï-Lama, Nil, Paris.
Varela F., Lachaux J.-P., Rodriguez E. & Mar-tinerie J. (2001) The brainweb : Phase syn-chronization and large-scale integration, Na-ture Reviews Neuroscience, 2, 229-239.
Varela F. & Maturana H. (1980) Autopoiesis andCognition : The Realization of the Living, Rei-del, Boston.
Varela F. & Shear J. (Eds). (1999) The Viewfrom Within : First-person Approaches to theStudy of Consciousness. London : Imprintacademic.
Varela F., Thomson E. & Rosch E. (1991) TheEmbodied Mind: Cognitive Science and Hu-man Experience, MIT Press; (1993) in French: L’inscription corporelle de l’Esprit. Sciencescognitives et Expérience humaine, Seuil,Paris.
Wallace R. K. (1970) Physiological effects oftranscendental meditation. Science, 167,1751-1754.
Wallace R. K. (1970) The physiological effectsof transcendental meditation, a proposed IVthmajor state of consciousness. Ph.D. Thesis,U.C.L.A., Los Angeles, 142 p.
Wallace R. K. & Benson H. (1972) The physiol-ogy of meditation. Scientific American, 226,84-90.
24 Human Cognitive Neurophysiology 2010, 3 (1)

P. Etevenon — ”Modified States of Consciousness” revisited
Wallace R. K., Benson H. & Wilson A. F. (1971)A wakeful hypometabolic physiologic state.Amer. J. Physiol., 221, 795-799.
Wallace R. K., Benson H. ,Wilson A. F. & GarrettM.D. (1971) Decreased blood lactate duringtranscendental meditation. Federation Pro-ceedings, 30, 376.
Walter G. (1953) The Living Brain, New York ;Penguin, London, 1967.
Walter D.O., Etevenon P., Pidoux B., Tortrat D.& Guillou S. (1984) Computerized topo-EEGspectral maps : difficulties and perspectives.Neuropsychobiology, 11, 264-272.
Wenger, M. A., & Bagchi, B. K. (1961). Studiesof autonomic functions in practitioners of yogain India. Behavioral Science, 6, 312–323.
Whyte L. (1959) The Unconscious Before Freud.Tavistock, London.
Zimmer H. (1953) Philosophies of India. Camp-bell J. (Ed.), Bollingen Series, Princeton.
2010, 3 (1) Kognitive Neurophysiologie des Menschen 25

K. Suzuki & H. Shinoda — Error-related Components and Impulsivity
Abstract
K. Suzuki & H. Shinoda (Tokyo, Japan) — Error-related Components and Impulsivity Related toSpeed and to Accuracy Trade-offWe examined the neurophysiological aspect of impulsivity as defined by performance. 71 participants
were classified into 3 subgroups by determining response time (RT) and rate of incorrect responses on
an arrow version of the flanker task: the impulsive group (+1SD above average or higher in incorrect
response rate), the average group (values in the range of mean ±1SD of RT and incorrect response rate),
and the remaining group. 7 participants from the impulsive (6 males and 1 females) and the average group
(5 males and 2 females) were investigated using error-related components. Compared to the average
group, a lower activity of error-related negativity (ERN/Ne) in the impulsive group was found as both the
amplitude and topographical differences. However no difference in error positivity (Pe) was seen in ether
group. These results suggest that an inability to adjust the behavior underlying impulsivity is associated
with the ERN/Ne activity.
Keywords: Error-related negativity or error negativity (ERN/Ne); Error positivity (Pe); Behavioral ad-
justment; Impulsivity
K. Suzuki & H. Shinoda —Error-related Componentsand Impulsivity related toSpeed and to Accuracy
Trade-off
K. Suzuki1 & H. Shinoda2
1Department of Psychology, 2Department ofClinical Psychology, Rissho University, Osaki
4-2-16, Shinagawa, Tokyo, [email protected]
Introduction
In response time (RT) tasks, participantssearch for an optimal RT band to achievethe appropriate balance between speed andaccuracy Rabbitt and Vyas (1970). If theyrespond faster than the optimal RT band(i.e. risky response), accuracy is decreased,whereas if they respond more slowly (i.e. cau-tion response), accuracy does not change.Individual differences in strategies betweenspeed and accuracy exist, and some partici-pants show a preference for risky response.Kagan (1965) defined the tendency to show apreference for a risky response as impulsivity.This definition of impulsivity was not asso-ciated with impulsivity measured by ques-tionnaire (Block, Block, & Harrington, 1974;
26 Human Cognitive Neurophysiology 2010, 3 (1)

K. Suzuki & H. Shinoda — Error-related Components and Impulsivity
Leshem & Glicksohn, 2007), but considereda separate dimension of impulsivity Dickman(1993); Leshem and Glicksohn (2007). Sinceself-monitoring their response is important forparticipants when searching for an optimal RTband, the neural system of monitoring behav-ior may operate differently in individuals withthe impulsivity related by speed and accuracytrade-off. The process of behavior monitor-ing can be investigated using error-relatednegativity or error negativity (ERN/Ne) anderror positivity (Pe) (Falkenstein, Hoormann,Christ, & Hohnsbein, 2000).
ERN/Ne is characterized by a negative de-flection in response-locked event related brainpotentials (ERPs) at the midline fronto-centralarea occurring during first 100ms following in-correct responses (Falkenstein et al., 2000;Gehring, Goss, Coles, Meyer, & Donchin,1993). Several theories have attempted to ex-plain the functional significance of ERN/Ne.Originally, ERN/Ne was thought to reflect theactivity of an error-detection system compar-ing a correct response to an actual response(Bernstein, Scheffers, & Coles, 1995). Morerecently, researchers proposed a link betweenerror detection and reinforcement learning inwhich ERN/Ne functions as a negative rein-forcement learning signal conveyed to the an-terior cingulate cortex (ACC) via the mesen-cephalic dopamine system (Holroyd & Coles,2002). Another proposal posits ERN/Ne aspart of a conflict monitoring system betweencorrect and incorrect responses. Accordingto this theory, ERN/Ne signals from the mon-itored conflict affect various control centersleading to top-down regulation for balancingresponses, e.g., between speed and accuracy(Botvinick, Braver, Barch, Carter, & Cohen,
2001).
Following after the ERN/Ne, Pe is observedas a positive deflection over parieto-centralareas (Falkenstein et al., 2000). Pe is as-sociated with conscious error recognitionand subsequent compensation (Nieuwenhuis,Ridderinkhof, Blom, Band, & Kok, 2001).
Although both ERN/Ne and Pe are involved inthe behavior monitoring process, the effectson adjustment of behavior are different. Inan anti-saccade task that requires a saccadeto the opposite side of a peripheral abruptonset cue, when participants make reflexiveand unperceived errors which could not becorrected on the next trial, Pe was substan-tially reduced in amplitude while ERN/Ne wasnot (Nieuwenhuis et al., 2001). Moreoversome studies reported that ERN/Ne was ob-served on correct trials (i.e., correct responsenegativity, CRN) but Pe was not (Bartholowet al., 2005; Vidal, Hasbroucq, Grapperon,& Bonnet, 2000). In contrast, the amplitudeof ERN/Ne was larger for trials judged ascertainly incorrect compared to trials judgedas ambiguous (Scheffers & Coles, 2000).In terms of personality traits, obsessive-compulsive disorder (OCD) patients, whohave negative emotions and engage in exces-sive self-monitoring, display larger ERN/Neamplitudes than control participants (Gehring,Himle, & Nisenson, 2000). These findingssuggest that Pe is specific to the error pro-cess and is directly associated with responsesfollowing incorrect trials, while ERN/Ne is notspecific to the error process but is associatedwith the value and negative emotions of theparticipants.
In the present study, we examined the rela-tionship between impulsivity as defined by
2010, 3 (1) Kognitive Neurophysiologie des Menschen 27

K. Suzuki & H. Shinoda — Error-related Components and Impulsivity
performance and the error-related ERP com-ponents. Kagan (1965) measured impulsivityutilizing a speed and accuracy trade-off ofperformance on a task in which participantswere asked to select the figure that exactlymatches the standard from the alternatives.Alternately, a flanker task (Eriksen & Erik-sen, 1979) has often been used to recorderror-related ERP components. Gehring etal. (1993) reported shortened RT and in-creased incorrect response rate in the flankertask with an increased demand for responsespeed. Hence we considered the individualdifferences of performance in the flanker taskwould reflect on the impulsivity.
Methods
Task
In an arrow version of the flanker task, partici-pants were required to press a button with thethumb of their left or right hand as quickly andcorrectly as possible, corresponding to the di-rection of the central arrow flanked by fourdistracter arrows. Stimulus consisted of twocompatible arrays, < < < < < and > > > > >.The probability of each these stimuli was .25.The stimulus was presented for 50 ms withstimuli onset asynchrony (SOA) of 2000 mson PC monitor using STIM2 software (Neu-roScan Inc.).
Participants and procedure
Participants were selected from a group of un-dergraduate and graduate students (42 malesand 29 females, age = 23.0 ± 2.9 years old).After giving informed consent, they performedthe flanker task before the main session. In
the recruiting session, compatible trials gavea correct RT of 367.07 ± 42.43 ms and an in-correct response rate of 0.53 ± 1.39%. Com-parable figure for incompatible trials gave acorrect RT of 424.41 ± 45.89 ms and an in-correct response rate of 4.77 ± 5.25%. Im-portantly, the individual difference was signifi-cantly larger for incompatible trials than com-patible trials (F (70) = 14.22, p <. 001). More-over we found a significant negative correla-tion between mean correct RT and incorrectresponse rate for the incompatible trials (r=-0.40, p <.001, Figure 1A). Thus we split par-ticipants into three subgroups based on +1SDof grand mean correct RT and incorrect re-sponse rate for incompatible trials (see Figure1A): The impulsive group (+1SD above aver-age or higher in incorrect response rate), theaverage group (values in the range of mean±1SD of RT and incorrect response rate), andthe remaining group. Agreeing to participatein the main session after the recruiting ses-sion were seven participants in the impulsivegroup (six males and one female, age = 22.3± 0.7 years old) and seven age-matched par-ticipants in the average groups (five males andtwo females, age = 22.5 ± 1.2 years old). Twoparticipants in the impulsive group describedthemselves as being left-handed. The recruit-ing session consisted of 3 blocks of 48 trialsafter a practice (48 trials) and the main ses-sion consisted of 16 blocks.
EEG recordings
The electroencephalograms (EEGs) wererecorded in 15 channels simultaneously froman array of electrodes located at F3, Fz,F4, T3, C3, Cz, C4, T4, T5, P3, Pz, P4,T6, O1 and O2. Vertical and horizontal
28 Human Cognitive Neurophysiology 2010, 3 (1)
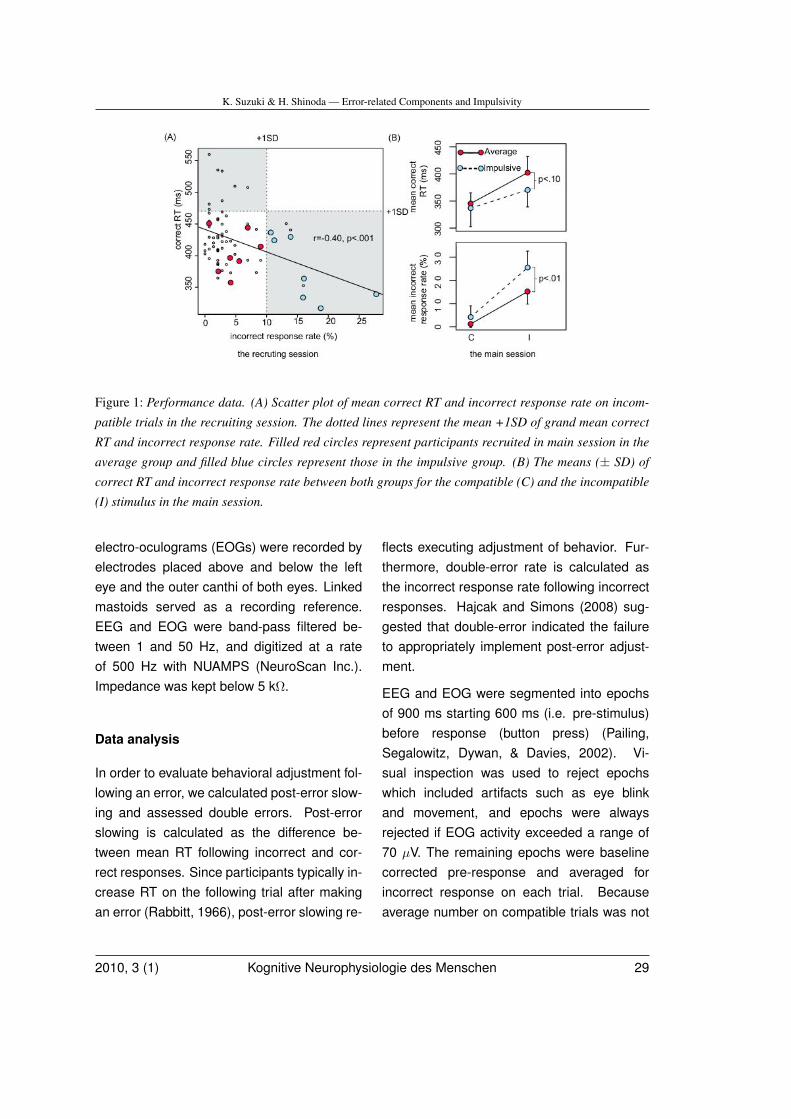
K. Suzuki & H. Shinoda — Error-related Components and Impulsivity
Figure 1: Performance data. (A) Scatter plot of mean correct RT and incorrect response rate on incom-
patible trials in the recruiting session. The dotted lines represent the mean +1SD of grand mean correct
RT and incorrect response rate. Filled red circles represent participants recruited in main session in the
average group and filled blue circles represent those in the impulsive group. (B) The means (± SD) of
correct RT and incorrect response rate between both groups for the compatible (C) and the incompatible
(I) stimulus in the main session.
electro-oculograms (EOGs) were recorded byelectrodes placed above and below the lefteye and the outer canthi of both eyes. Linkedmastoids served as a recording reference.EEG and EOG were band-pass filtered be-tween 1 and 50 Hz, and digitized at a rateof 500 Hz with NUAMPS (NeuroScan Inc.).Impedance was kept below 5 kΩ.
Data analysis
In order to evaluate behavioral adjustment fol-lowing an error, we calculated post-error slow-ing and assessed double errors. Post-errorslowing is calculated as the difference be-tween mean RT following incorrect and cor-rect responses. Since participants typically in-crease RT on the following trial after makingan error (Rabbitt, 1966), post-error slowing re-
flects executing adjustment of behavior. Fur-thermore, double-error rate is calculated asthe incorrect response rate following incorrectresponses. Hajcak and Simons (2008) sug-gested that double-error indicated the failureto appropriately implement post-error adjust-ment.
EEG and EOG were segmented into epochsof 900 ms starting 600 ms (i.e. pre-stimulus)before response (button press) (Pailing,Segalowitz, Dywan, & Davies, 2002). Vi-sual inspection was used to reject epochswhich included artifacts such as eye blinkand movement, and epochs were alwaysrejected if EOG activity exceeded a range of70 µV. The remaining epochs were baselinecorrected pre-response and averaged forincorrect response on each trial. Becauseaverage number on compatible trials was not
2010, 3 (1) Kognitive Neurophysiologie des Menschen 29

K. Suzuki & H. Shinoda — Error-related Components and Impulsivity
enough, event-related potentials (ERPs) forcompatible trial were excluded from analyses.In order to focus on the temporal dynamicsof potential field strength and topographicaldistribution, we analyzed GFP and centroidlocations (positive or negative locations in theanterior-posterior or in the left-right direction)at each time point using average reference(Lehmann & Skrandies, 1980; Skrandies,1987). GFP was computed as the spatialSD of overall voltages in each map and itspeaks represents times of maximum overallfield strength corresponding to latencies ofthe major ERP components. To examine thespatial characteristics of the brain electricfield, the centroid locations were computed asthe scalp position of the centers of gravity inthe positive and negative map area.
Results
Behavioral data
Figure 1B shows the mean (± SD) for correctRT and incorrect response rate. We observedthat individuals in the impulsive group hadshorter correct RT and higher incorrect re-sponse rate compared to individuals in the av-erage group. The difference was remarkablefor incompatible trials. To analyze correct RTand incorrect response rate, we submitted toseparate repeated measures analyses of vari-ance (ANOVAs) with 2 groups × 2 compatibil-ity as factors. We found a significant compati-bility effect (F (1, 12) = 140.74, p <. 01) and asignificant interaction of groups by compatibil-ity (F (1, 12) = 9.45, p <. 01). The method ofHolm (Holm, 1979) was used to adjust the Pvalues in multiple testing. Holm tests revealedthat compatible trials were shorter than incom-
patible trials in both groups (p <. 01). In addi-tion, individuals tended to have shorter correctRT in the impulsive than the average group forincompatible trials (p <. 10). However, a sig-nificant group effect was not found (F (1, 12) =1.79, p = .20). In terms of incorrect responserate, we found a significant compatibility ef-fect (F (1, 12) = 114.10, p <. 01), a significantgroup effect (F (1, 12) = 9.16, p <. 05), a sig-nificant interaction of groups by compatibility(F (1, 12) = 5.00, p <. 05). Holm tests revealedthat individuals had higher incorrect responserate in the impulsive than the average groupfor incompatible trials (p <. 01) and compat-ible trials showed a lower incorrect responserate than incompatible trials in both groups (p<. 01).
As to indices of behavioral adjustment, twosamples t-tests were performed, and showedfollowing results. Double-error rate was higherin the impulsive than the average group (t(12)= -3.25, p <. 01; average group = 6.67 ±5.27%, impulsive group = 16.53 ± 6.03%).However, both groups did not differ in post-error slowing (t(12) = 1.25, p = .23; averagegroup = 8.85 ± 10.75 ms, impulsive group =2.83 ± 13.76 ms).
Electrophysiological data
Response-locked ERPs and GFP peaks
Figure 2A shows the grand mean response-locked ERP waveforms and GFP peaksevoked by incorrect responses in incompat-ible trials. Clear GFP peaks can be seenat 24 ms for the first component and at 156ms for the second component in the averagegroup and at 22 ms for the first componentand at 138 ms for the second component in
30 Human Cognitive Neurophysiology 2010, 3 (1)

K. Suzuki & H. Shinoda — Error-related Components and Impulsivity
Figure 2: Grand mean response-locked ERP waveforms on incorrect trials for the average and impulsive
individuals. (A) ERP waveforms and GFP, (B) ERP waveforms at electrode site Cz, (C) Mean amplitudes
of ERN/Ne and Pe at electrode site Cz between both groups.
the impulsive group (Figure 2A, low). Thesecomponents were identified as ERN/Ne andPe. We compared difference of the meanGFP value and amplitude at Cz (Figure 2B) ofERN/Ne and Pe between both groups by twosample t-test and Welch’s t-test. No significantdifference was found between either group forthe GFP value of ERN/Ne or Pe (t(12) = 0.92,p = .38; t(7.6) = 0.76, p = .47). For the ampli-tude of ERN/Ne and Pe (Figure 2B), ANOVAswith 2 groups × 3 electrodes (Fz, Cz, Pz)using Greenhouse-Geisser correction wereperformed. There was a significant interactionof groups by electrodes in the amplitude ofERN/Ne (F (1,12) = 28.07, p <. 05, ε = 0.63).Holms tests revealed that the difference ofthe amplitude of ERN/Ne at Cz (p <. 05)and a tendency level at Pz (p = .05). On theother hand, there were not a significant maineffect of groups and a significant interaction
of groups by electrodes in the amplitude of Pe(F (1,12) = 0.46, p = .50, ε = 0.67; F (1,12) =0.98, p = .36, ε = 0.67).
Correlational findings
To investigate the relationship between ERPcomponents and the number of errors, we per-formed correlational analyses between ERPcomponents and incorrect response rate onincompatible trials. A tendency was found forthe amplitude of ERN/Ne at Cz (r = 0.48, p <.08), while there was not any significant corre-lation with the amplitude of Pe at Cz(r =-0.35,p = .22). Furthermore, we analyzed the aver-age and impulsive groups separately. For theimpulsive group, we found a significant corre-lation with the amplitude of ERN/Ne at Cz (r =0.83, p <. 05), whereas we could not find anysignificant correlation for the average group (r= -0.35, p = .43). Neither was there any signif-
2010, 3 (1) Kognitive Neurophysiologie des Menschen 31

K. Suzuki & H. Shinoda — Error-related Components and Impulsivity
icant correlation with the amplitude of Pe at Czfor the average group nor the impulsive group(r = -0.30, p = .49; r = -0.08, p = .85).
Topographical analysisFigure 3A shows an ERP map series due toincorrect responses on incompatible trials. Ascan be seen, the change in ERP topographyoccurred between 0 and 40ms after the re-sponse. To investigate this difference (the oc-currence of ERN/Ne) in detail, the centroid lo-cation was calculated. The trajectory of nega-tive and positive centroids is illustrated in Fig-ure 3B. Although the location of negative cen-troids in the average group moved from theanterior to the posterior, those of the impul-sive group remained in the anterior area. Thetwo samples t-test shows that the location ofnegative centroid in the Y direction (i.e. an-terior from posterior) at ERN/Ne peak in theimpulsive group distributed significantly moreanterior than that in the average group (t(12)= 2.28, p <. 05).
Discussion
In the present study, participants who had ashorter mean correct RT tended to have ahigher incorrect response rate while partic-ipants who had a longer mean correct RTtended to have a lower incorrect responserate in the flanker task. Based on this re-lationship between mean correct RT and in-correct response rate (i.e. speed and accu-racy trade-off), we assessed impulsivity as thetendency to commit more errors and respondmore quickly. In terms of behavioral adjust-ment, although post-error slowing did not sig-nificantly differ in both groups, these resultssuggest that individuals in the impulsive group
needed to respond more slowly, as RT in theimpulsive group tended to be faster than in theaverage group. Accordingly, the double-errorrate was higher in the impulsive group whencompared to the average group. A previousstudy reported that double-error indicated fail-ure to appropriately implement post-error ad-justment (Hajcak & Simons, 2008). Thus, in-dividuals in the impulsive group may not ad-just response adequately after incorrect re-sponses.
More importantly, our findings indicated thatthis inability to adjust behavior in the impulsivegroup was associated with a decrease in theamplitude of the ERN/Ne component. Currentcomputational accounts of the role of ERN/Nein reinforcement learning theory (Holroyd &Coles, 2002) and conflict monitoring theory(Botvinick et al., 2001) predicted these re-sults. These models suggest that an error ora conflict signal reflected by ERN/Ne leads tobehavioral adjustment; empirical studies alsoshow support for these results (Debener etal., 2005; Gehring et al., 1993). A studyexploring single-trial ERN/Ne using indepen-dent component analysis reported that longerRT on subsequent trials were associated withhigher amplitudes of ERN/Ne (Debener et al.,2005). Moreover, earlier studies showed thatERN/Ne amplitude covaried with perceived in-correct responses by participants (Scheffers &Coles, 2000). These findings suggest that in-dividuals in the impulsive group are unsuscep-tible to the value of incorrect response.
In scalp field topography, we found that thenegative centroids at ERN/Ne peak was cen-tral area in the average group while it wasmore anterior area in the impulsive group,although there was not a significant differ-
32 Human Cognitive Neurophysiology 2010, 3 (1)
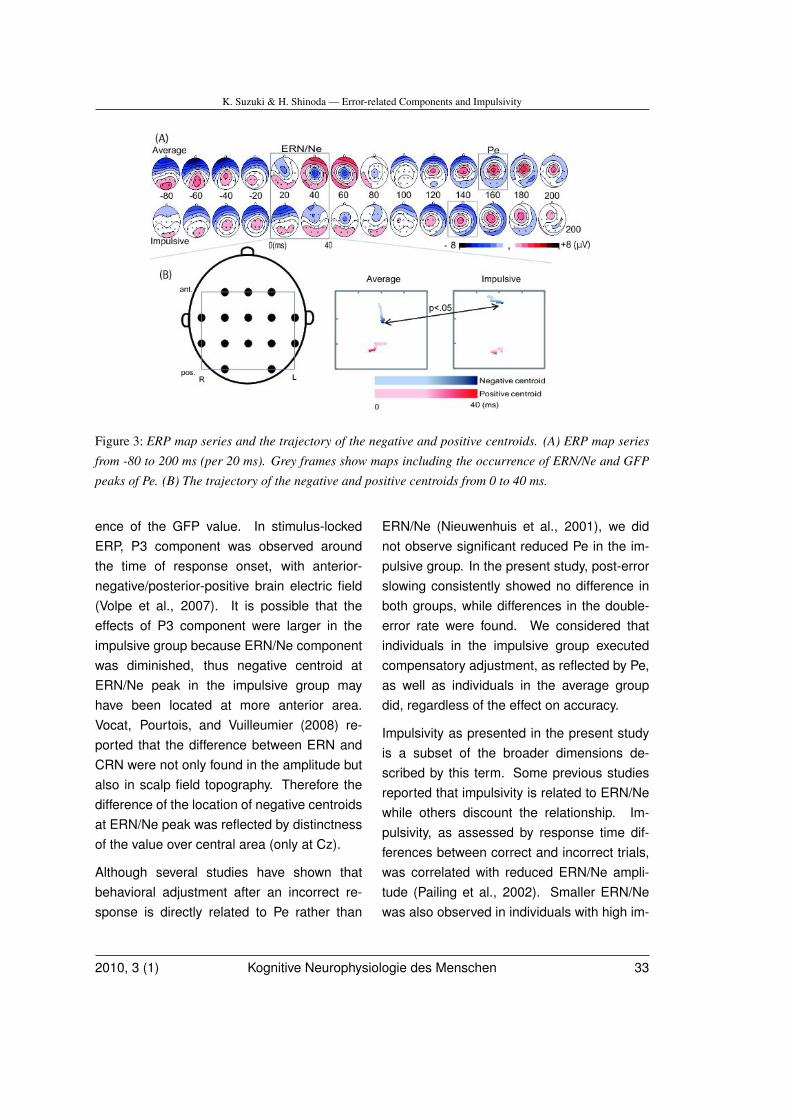
K. Suzuki & H. Shinoda — Error-related Components and Impulsivity
Figure 3: ERP map series and the trajectory of the negative and positive centroids. (A) ERP map series
from -80 to 200 ms (per 20 ms). Grey frames show maps including the occurrence of ERN/Ne and GFP
peaks of Pe. (B) The trajectory of the negative and positive centroids from 0 to 40 ms.
ence of the GFP value. In stimulus-lockedERP, P3 component was observed aroundthe time of response onset, with anterior-negative/posterior-positive brain electric field(Volpe et al., 2007). It is possible that theeffects of P3 component were larger in theimpulsive group because ERN/Ne componentwas diminished, thus negative centroid atERN/Ne peak in the impulsive group mayhave been located at more anterior area.Vocat, Pourtois, and Vuilleumier (2008) re-ported that the difference between ERN andCRN were not only found in the amplitude butalso in scalp field topography. Therefore thedifference of the location of negative centroidsat ERN/Ne peak was reflected by distinctnessof the value over central area (only at Cz).
Although several studies have shown thatbehavioral adjustment after an incorrect re-sponse is directly related to Pe rather than
ERN/Ne (Nieuwenhuis et al., 2001), we didnot observe significant reduced Pe in the im-pulsive group. In the present study, post-errorslowing consistently showed no difference inboth groups, while differences in the double-error rate were found. We considered thatindividuals in the impulsive group executedcompensatory adjustment, as reflected by Pe,as well as individuals in the average groupdid, regardless of the effect on accuracy.
Impulsivity as presented in the present studyis a subset of the broader dimensions de-scribed by this term. Some previous studiesreported that impulsivity is related to ERN/Newhile others discount the relationship. Im-pulsivity, as assessed by response time dif-ferences between correct and incorrect trials,was correlated with reduced ERN/Ne ampli-tude (Pailing et al., 2002). Smaller ERN/Newas also observed in individuals with high im-
2010, 3 (1) Kognitive Neurophysiologie des Menschen 33

K. Suzuki & H. Shinoda — Error-related Components and Impulsivity
pulsivity related to performance of the stopsignal task and personality traits (Stahl & Gib-bons, 2007). However, the ranking of im-pulsiveness as one of the personality traitswas not related to ERN/Ne (Santesso & Sega-lowitz, 2009). According to previous studies,at least, the impulsivity associated with taskperformance may be related to ERN/Ne.
The impulsive group had a higher incorrectresponse rate than the average group, so itis possible that the reduced ERN/Ne in theimpulsive group reflects simple habituation.However, we believe that this likely is not thecase for the following reason. If simple habitu-ation affected ERN/Ne, there would be a pos-itive correlation between ERN/Ne amplitudeand incorrect response rate in both groups.Although a significant positive correlation wasobserved in the impulsive group, no signifi-cant correlation could be found in the averagegroup (furthermore negative correlation wasobserved). We speculate that ERN/Ne wasaffected by response style rather than any ha-bituation effect.
The present study had several limitations.First, the small sample size may be problem-atic. It suggests limited statistical power to de-tect statistical significance. Furthermore, theaverage and impulsive groups was not con-trolled for gender and handedness. Althoughthe effects of gender and handedness mightaffect ERN/Ne components, we could not con-firm them statistically because of the sam-ple size. Future studies should use largersample size in order to clarify these issues.Another limitation related to number of elec-trodes. In general, the amplitude of ERN/Newas maximal at FCz and Pe was maximal atCPz (Falkenstein et al., 2000). However these
channels were not recorded in the presentstudy. Therefore, future studies should inves-tigate GFP value and the locations of negativeand positive centroids due to ERN/Ne and Peusing higher-density EEG recordings. Finally,although the impulsivity was defined with RTand incorrect response rate, we recruited par-ticipants in the impulsive group based on onlyincorrect response rate. Thus some of themhad longer correct RT than some of partic-ipants in the average group. Moreover, theability to engage in the flanker task is differentin participants (e.g. the variety of incorrect re-sponse rate over participants who had samecorrect RT). These facts suggest the defini-tion of impulsivity is ambiguous in the presentstudy. However, in the view of speed and ac-curacy trade-off within a participant, a highincorrect response rate shows that responsetimes were short for the participant (Rabbitt &Vyas, 1970). Thus, it might be appropriate torecruit the participants with incorrect responserate in the present study.
In summary, performance data (correct RTand double-error rate) indicates an inability bythe impulsive group to adjust behavior. Dif-ferences in ERN/Ne activity in the impulsivegroup is seen in both lowered amplitude andtopographical difference. These results sug-gest that the inability to adjust behavior in theimpulsive group is related to relatively low val-ues of ERN/Ne to incorrect responses.
Acknowledgements
We would like to thank Prof. Wolfgang Skran-dies for thoughtful suggestions.
34 Human Cognitive Neurophysiology 2010, 3 (1)

K. Suzuki & H. Shinoda — Error-related Components and Impulsivity
References
Bartholow, B. D., Pearson, M. A., Dickter,C. L., Sher, K. J., Fabiani, M., & Gratton,G. (2005). Strategic control and medialfrontal negativity: Beyond errors and re-sponse conflict. Psychophysiology , 42,33–42.
Bernstein, P. S., Scheffers, M. K., & Coles,M. G. H. (1995). ”Where Did I GoWrong?” A psychophysiological analy-sis of error detection. Journal of Exper-imental Psychology: Human Perceptionand Performance, 21, 1312–1322.
Block, J., Block, J. H., & Harrington, D. M.(1974). Some misgivings about theMatching Familiar Figures Test as ameasure of reflection-impulsivity. Devel-opmental Psychology , 10, 611–632.
Botvinick, M. M., Braver, T. S., Barch, D. M.,Carter, C. S., & Cohen, J. D. (2001).Conflict monitoring and cognitive con-trol. Psychological Review , 108, 624–652.
Debener, S., Ullsperger, M., Siegel, M.,Fiehler, K., Von Cramon, D. Y., & Engel,A. K. (2005). Trial-by-trial coupling ofconcurrent electroencephalogram andfunctional magnetic resonance imagingidentifies the dynamics of performancemonitoring. Journal of Neuroscience,25, 11730–11737.
Dickman, S. J. (1993). Impulsivity and infor-mation processing. In W. G. McCown,J. L. Johnson, & M. B. Shure (Eds.), Theimpulsive client: Theory, research, andtreatment (pp. 151–184). Washington,D.C.: American Psychological Associa-tion.
Eriksen, C. W., & Eriksen, B. A. (1979). Tar-get redundancy in visual search- Do rep-etitions of the target within the displayimpair processing. Perception and Psy-chophysics, 26, 195–205.
Falkenstein, M., Hoormann, J., Christ, S., &Hohnsbein, J. (2000). ERP componentson reaction errors and their functionalsignificance: a tutorial. Biological Psy-chology , 51, 87–107.
Gehring, W. J., Goss, B., Coles, M. G. H.,Meyer, D. E., & Donchin, E. (1993).A neural system for error detection andcompensation. Psychological Science,4, 385–390.
Gehring, W. J., Himle, J., & Nisenson, L. G.(2000). Action-monitoring dysfunction inobsessive-compulsive disorder. Psycho-logical Science, 11, 1–6.
Hajcak, G., & Simons, R. F. (2008). Oops!.. I did it again: An ERP and behavioralstudy of double-errors. Brain and Cog-nition, 68, 15–21.
Holm, S. (1979). A simple sequentially rejec-tive multiple test procedure. Scandina-vian Journal of Statistics, 6, 65–70.
Holroyd, C. B., & Coles, M. G. H. (2002). Theneural basis of human error processing:reinforcement learning, dopamine, andthe error-related negativity. Psychologi-cal Review , 109, 679–708.
Kagan, J. (1965). Individual differences in theresolution of response uncertainty. Jour-nal of Personality and Social Psychol-ogy , 2, 154–160.
Lehmann, D., & Skrandies, W. (1980).Reference-free identification of com-ponents of checkerboard-evoked mul-tichannel potential fields. Electroen-
2010, 3 (1) Kognitive Neurophysiologie des Menschen 35

K. Suzuki & H. Shinoda — Error-related Components and Impulsivity
cephalography and Clinical Neurophys-iology , 48, 609–621.
Leshem, R., & Glicksohn, J. (2007). The con-struct of impulsivity revisited. Personal-ity and Individual Differences, 43, 681–691.
Nieuwenhuis, S., Ridderinkhof, K. R., Blom,J., Band, G. P. H., & Kok, A. (2001).Error-related brain potentials are dif-ferentially related to awareness of re-sponse errors: Evidence from an anti-saccade task. Psychophysiology , 38,752–760.
Pailing, P. E., Segalowitz, S. J., Dywan, J.,& Davies, P. L. (2002). Error negativ-ity and response control. Psychophysi-ology , 39, 198–206.
Rabbitt, P. (1966). Errors and error correctionin choice-response tasks. Journal of Ex-perimental Psychology , 71, 264–272.
Rabbitt, P., & Vyas, S. (1970). An elementarypreliminary taxonomy for some errors inlaboratory choice RT tasks. Acta Psy-chologica, 33, 56–76.
Santesso, D. L., & Segalowitz, S. J. (2009).The error-related negativity is related torisk taking and empathy in young men.Psychophysiology , 46, 143–152.
Scheffers, M. K., & Coles, M. G. H. (2000).Performance monitoring in a confusingworld: Error-related brain activity, judg-ments of response accuracy, and typesof errors. Journal of Experimental Psy-chology: Human Perception and Perfor-mance, 26, 141–151.
Skrandies, W. (1987). The upper and lowervisual field of man: Electrophysiologicaland functional differences. Progress insensory physiology , 8, 1–93.
Stahl, J., & Gibbons, H. (2007). Dynam-ics of response-conflict monitoring andindividual differences in response con-trol and behavioral control: An electro-physiological investigation using a stop-signal task. Clinical Neurophysiology ,118, 581–596.
Vidal, F., Hasbroucq, T., Grapperon, J., & Bon-net, M. (2000). Is the ”error negativity”specific to errors? Biological Psychol-ogy , 51, 109–128.
Vocat, R., Pourtois, G., & Vuilleumier, P.(2008). Unavoidable errors: a spatio-temporal analysis of time-course andneural sources of evoked potentials as-sociated with error processing in aspeeded task. Neuropsychologia, 46,2545–2555.
Volpe, U., Mucci, A., Bucci, P., Merlotti, E.,Galderisi, S., & Maj, M. (2007). Thecortical generators of P3a and P3b: ALORETA study. Brain Research Bulletin,73, 220–230.
36 Human Cognitive Neurophysiology 2010, 3 (1)

W. Skrandies — Abstracts of the 18th German EEG/EP Mapping Meeting
Abstracts of the 18thGermanEEG/EP Mapping Meeting,Giessen, October 16 - 18,
2009
Altered prefrontal activation duringdecision-making in panic patients: im-pact of error feedback in previous trial.L.H. Ernst (1), A.-C. Ehlis (1), M.M. Plichta(1), T. Dresler (1), S. Tupak (1), B. War-rings (1), P. Pauli (2) & A.J. Fallgatter (1),(1)Psychophysiology and Functional Imaging,Department of Psychiatry, Psychosomaticsand Psychotherapy, University of Würzburg,Würzburg, Germany; (2) Biological Psychol-ogy, Clinical Psychology and Psychotherapy,University of Würzburg, Würzburg, GermanyPrevious research in decision-making couldshow that panic patients are more likely thanhealthy persons to change their behaviouralstrategy after the occurrence of an error. Sucha pattern was interpreted as a permanentsearch for an optimal decision-making strat-egy in panic patients. The assumed anatomi-cal basis of this process is a dysfunctional in-teraction between the anterior cingulate cor-tex (ACC) - as a monitoring system for er-rors - and the dorsolateral prefrontal cortex(dlPFC) - as the main regulation instance indecision-making. We investigated the role ofthe dlPFC during a simple decision-makingtask with a predefined error probability in agroup of panic patients and healthy controls.Groups were compared in regard to strat-egy choices and brain activity during decisionmaking for the two conditions ’error feedbackin the previous trial’ and ’positive feedback in
the previous trial’. Brain activity was mea-sured with near-infrared spectroscopy (NIRS)and electroencephalography (EEG); addition-ally, the skin conductance response (SCR)was recorded. Preliminary results suggestaltered activation patterns in the dlPFC dur-ing decision-making in panic patients in com-parison to healthy controls following an er-ror feedback in the previous trial. Both spa-tial and temporal activation patterns show sig-nificant aberrances. Therefore, the assump-tion of dysfunctional dlPFC processes duringdecision-making in panic patients can be sup-ported. Especially because of the altered tem-poral process, our result further underpins thenotion of a modified influence of error-relatedinformation on subsequent decision making.Results are discussed regarding the role of al-tered error-processing in the maintenance ofsafety seeking behaviour and thereby of panicdisorder.
Frontal midline theta reflecting workingmemory load: Influence of selected fre-quencies and of variability in behavioraleffects.U. Maurer, S. Brem, M. Liechti, S. Maurizio, R.Lüchinger, K. Bucher, L. Michels & D. Bran-deis, Zentrum für Kinder und Jugendpsychia-trie, Universität Zürich, Zürich, Switzerland.Frontal midline theta (4-8Hz) activity has beenrelated to working memory (WM) processes,as it typically increases with WM load. The ro-bustness of this effect, however, varies acrossstudies and subjects, posing limits to its in-terpretation. We hypothesized that the frontalmidline theta WM effect is not only topograph-ically focal at anterior-frontal electrodes, butmay also occur at a very narrow and individ-ually variable frequency band. In addition, we
2010, 3 (1) Kognitive Neurophysiologie des Menschen 37

W. Skrandies — Abstracts of the 18th German EEG/EP Mapping Meeting
hypothesized that individual variability in thebehavioral WM effect may explain some vari-ation in the frontal midline theta WM effect.We recorded a 63-channel EEG from healthy,young adults while they performed a visualWM task with unfamiliar symbols under low(2 items) and high (4 items) workload con-ditions. Preliminary results from the first 15subjects show that higher workload led to lessaccurate and slower responses at the behav-ioral level (accuracy: 83.3 vs. 93.8%; reac-tion time: 1132 vs. 1000ms). While frontalmidline theta increased with workload, this in-crease was only robust using individually de-termined theta peaks (AFz: 58.8%; t=4.09,p<0.01), but not for the mean theta band (4-8Hz; AFz: 10.6%; t=1.57, p=ns). In addition,the workload-dependent increase of theta ac-tivity was associated with a corresponding de-crease in accuracy (individual peak: r=-0.52,p<0.05). This correlation also held, if onlytrials with correct responses were includedin the EEG analysis, although only at trendlevel (r=-0.45, p<0.1). The results indicatethat workload-related theta effects are not onlytopographically focused around frontal mid-line electrodes, but are also limited to nar-row, individual theta frequency bands. Thestudy also shows correlations between behav-ioral responses and changes in frontal mid-line theta, and thus strengthens the notion thatfrontal midline theta reflects active WM pro-cesses.
EEG parameter seen with successful golfputts.M. Doppelmayr (1), T. Finkenzeller (2) & G.Amesberger (2), (1) Department of Physiolog-ical Psychology, University of Salzburg, Aus-tria (2) Department of Sport Science & Kine-
siology University of Salzburg, Austria.
Continuous attention to a stimulus usuallyevokes a specific kind of cortical activitytermed frontal midline theta (Fmθ). This Fmθcan be observed in the electroencephalogram(EEG) most pronounced at the frontal midlineelectrode location Fz and has been linked toseveral attention related processes, as actionregulation, concentration, sustained and in-ternalized attention. In general, it has beenrepeatedly shown that Fmθ is clearly linkedto the amount of mental effort and attention.The fact that Fmθ is related to attentionalprocesses led to the hypothesis, that thesedifferences in the allocation of attention re-lated resources, should be observable duringthe preparatory phase of a golf putt. 15 maleparticipants with different experience in golfperformed 100 putts (indoor) while multichan-nel EEG was recorded. For each putt thedistance to the target position was assessed.According to the average performance thesample was split in a group of good anda group of bad golfer. The putts of eachparticipant were divided successful and lesssuccessful trials. Fmθ activity was analyzedfor four 500 ms time intervals (- 1500 to + 500)around the backswing of the putt. A three fac-torial ANOVA for TIME X PERFORMANCE XGROUP calculated for Fz, revealed significantresults for TIME, PERFORMANCE, and TIMEX GROUP. Fmθ power is most pronounced atabout 1.5 sec before the swing of the putt anddecreases towards the putt. Interestingly, thiseffect is significantly stronger for good golfersas compared to those with less experience.Finally more successful putts are precededby a higher Fmθ power. Additional analysesindicated that these effects are reflected only
38 Human Cognitive Neurophysiology 2010, 3 (1)

W. Skrandies — Abstracts of the 18th German EEG/EP Mapping Meeting
at frontal electrode sites. Summarizing theseresults indicate that successful golf putts arepreceded by higher Fmθ indicating more fo-cused attention. These attentional processesare more pronounced in experienced golfers.
The functional meaning of human alphaoscillations for conscious perception.R. Freunberger (1,2) & W. Klimesch (1),(1) Department of Psychology, University ofSalzburg, Austria (2) Max-Planck Institute forHuman Development, Berlin, Germany.The amplitude increase of the human alpharhythm in an experimental situation has for along time been considered reflecting an idlingprocess without any functional meaning. Re-cent studies show that not only the alpha de-crease but also the alpha amplitude increaseserve specific cognitive functioning, mainlythe inhibition of task irrelevant brain areas orthe inhibition of distracting information. In ad-dition also the alpha phase seems to be im-portant for the exact timing of neural activityto establish efficient perception performance.In one of our studies images were presentedwith different degradation levels and subjectswere asked to indicate whether the presentedimage was a real or a nonsense object. Theresults show that within the time-window ofobject recognition alpha amplitude decreasesstronger for real as compared to nonsense ob-jects. Additionally, we found a stronger al-pha network for real objects within this timewindow as derived from phase-locking analy-ses. We interpret this alpha-network as an im-portant top-down mechanism that establishesthe access to semantic long-term memory.A follow-up study was designed using video-sequences with the same task manipulationas in the first experiment. But, in this task, we
did not have any ëventsänd, thus, can excludeany effects elicited by evoked activity. Prelim-inary data indicates that we can replicate thefindings from the first study and we can ex-tend the findings by showing that alpha is es-pecially relevant for the processing of mean-ingful information when compared to simpleprocessing of physical stimulus features. Weconclude that the onset of alpha desynchro-nization reflects the time point of successfulaccess to semantic long-term memory traces.
EEG LORETA lagged coherence duringresting, attention to calculation and atten-tion to breathing.P. Milz, P.L. Faber, R.D. Pascual-Marqui & D.Lehmann, The KEY Institute for Brain-MindResearch, University Hospital of Psychiatry,Zurich, Switzerland.Do different types of attention show relatedtendencies of intracerebral functional con-nectivity? 58-channel EEG of 25 healthy,meditation-naïve, right-handed, male stu-dents was recorded during three conditionsof 5 minutes each with closed eyes in ran-domized order: (1) resting [3 runs], (2) mentalarithmetic [2 runs], and (3) breath counting,a meditation initiation technique [2 runs]. Ar-tifact]controlled EEG data were recomputed,for the 8 EEG frequency bands, into eLORETAintracerebral current densities. Interpretationof functional connectivity between brain areaswhen using head surface]recorded EEG suf-fers from (1) uncertainty about intracerebralsource localization and (2) effects of volumeconduction. Computing coherence via sourcemodeling (e.g. LORETA) avoids problem (1),and excluding coherences with zero phase lagavoids problem (2). Accordingly, we computed”lagged coherence” intracerebral connectivity
2010, 3 (1) Kognitive Neurophysiologie des Menschen 39

W. Skrandies — Abstracts of the 18th German EEG/EP Mapping Meeting
between eLORETA current densities in 19 re-gions of interest. Averaged resting conditionwas compared to the breath counting and thearithmetic run of which participants post-hocreported that their concentration during taskperformance was best. Paired t-tests betweenconditions yielded differences of coherenceat p<0.1 (corrected for multiple testing) for4 frequency bands (alpha1, beta2, beta3,gamma) [number of connections in brackets]:breath counting showed lower coherencethan rest (all 4 bands [2,1,1,6]) and arithmetic(alpha1, beta2, gamma [1,2,2]); arithmeticshowed higher coherence than rest (gamma[1]). In sum, of all three conditions, breathcounting showed lowest demands on intrac-erebral functional connectivity. The results donot support generally increased coherenceduring attention. Instead, they suggest thatbreath counting and mental arithmetic inducemental states whose inter-area connectivitydiffers in several frequency bands. In thegamma frequency band, their inter-area con-nectivity differs in opposite directions fromrest.
Special features of intraoperative monitor-ing during selective dorsal rhizotomy.S. Wolter (1), C. Spies (1), C. Wagner (2),T. Michael (2), M. Schulz (3) & E.J. Haberl(3), (1) Anesthesiology and Operative Inten-sive Care Medicine; (2) Pediatric Neurology;(3) Pediatric Neurosurgery of the Charité -Universitätsmedizin Berlin, 13353 Berlin, Ger-many.In a selective dorsal rhizotomy (SDR), partialdeafferentiation of specifically targeted sen-sory nerve roots is carried out through a min-imal monosegmental inlet located above theconus medullaris. The decision as to which
parts of the roots are to be transected is notleft to chance, but is rather determined inthe course of intraoperative neurophysiolog-ical monitoring (IONM). The following proce-dure is used for each side of the body, for ev-ery segmental level (L2 to S1), and for everyrootlet: First the reflex threshold intensity isdetermined through repetitive single stimula-tion of the sensory nerve root, during whichthe response potential of the relevant mus-cle groups turns up on the electromyogram(EMG). A one-second 50Hz-train-stimulationis then administered in accordance with thedetermined intensity of reflex threshold. Sub-sequently the 50 Hz response is graded inkeeping with precisely defined criteria rangingfrom 0 to 4. On the basis of this grading andthe specific pattern involved, a partial sectionof 50 percent per nerve root is always per-formed. The stimulus responses in childrenwith infantile cerebral palsy who suffer motordisturbances as a result of their disorder varyfrom case to case and often deviate from thephysiological responses one would normallyexpect. The peculiarities which were at oddswith basic rules of anatomy and physiology in-volve the following:
- spreading of segmental innervation into re-spective muscle groups
- intensity required in order to reach reflexthreshold in dorsal nerve roots
- assumption that a right or left stimulation canresult in reflex responses only in the relevantipsilateral muscle groups.
In particular the special features of IONM willbe elaborated upon. SDR is a microsurgi-cal intervention. After more than 50 rhizo-tomies carried out at the Charité Hospital inthe above-described manner, a striking reduc-
40 Human Cognitive Neurophysiology 2010, 3 (1)

W. Skrandies — Abstracts of the 18th German EEG/EP Mapping Meeting
tion in spasticity was observed, along withsubstantial improvement in children’s ability towalk.TS Park and JM Johston: Surgical techniques ... Neuro-
surg. Focus, 2006, 21, E7 (1-6 ).
Evoked and induced event-related EEG os-cillations in high-performing young and el-derly individuals during a cognitive flexi-bility task.B. Kopp, Ch. Uhlmann, L. Behrmann, J. Howe& K. Wessel, Institut für Kognitive Neurolo-gie an der Technischen Universität Carolo-Wilhelmina zu Braunschweig und Neurologis-che Klinik, Klinikum Braunschweig, Germany.We conducted a study of normal brain agingthat was based upon the quantitative analysisof event-related EEG biosignals. Forty partici-pants (twenty-four young, mean age 22 years;sixteen elderly, mean age 70 years) tookpart in a task that was designed to challengecognitive flexibility. Elderly participants hadslower RTs than young participants, whereaserror rates did not differ between the two agegroups. The analysis of evoked and inducedevent-related EEG oscillations rested onMorlet wavelet transformations in the delta-,theta-, alpha- and beta-frequency bands. Theanalysis of the evoked event-related EEGoscillations revealed more pronounced cen-tral delta and theta activity in response to anauditory stimulus in the young compared tothe elderly participants. We also observeda crossover interaction with regard to theinduced EEG oscillations. Specifically, youngparticipants showed more pronounced event-related alpha-desynchronization at temporo-parietal electrodes during stimulus recogni-tion, whereas elderly participants showedevidence of more pronounced event-related
beta-desynchronization at central electrodesduring response preparation. These data arediscussed against the background of thalamo-cortical mechanisms of the genesis of EEGrhythms. Specifically, process-induced fa-cilitation of posterior thalamo-cortical pro-cessing (stimulus recognition) and of anteriorbasal ganglia-thalamo-cortical processing(response preparation) seem to be age-dependent. Future studies will examine therole of dopamine in these EEG phenomenathat are related to the aging of the brain.
Age-related feedback effects on workingmemory performance and brain activity.S. A. Schapkin & G. Freude, Federal Insti-tute of Occupational Safety and Health, Berlin,Germany.Working memory (WM) performance declineswith ageing due to significant neural decline.The aging process can be counteracted byreorganizing brain functions and changing ofcognitive strategies in older people. Feedback(FB) is assumed to help chunking informationand facilitating WM performance. Twenty oneyounger and 21 older employees had to per-form a visual 0-back (low WM load -WML) and2-back task (high WML) with and without FB.Under high WML older adults responded moreslowly and less accurately than younger ones.The P3b component was shifted to frontal ar-eas in older adults, while a parietal maximumin younger was found. P3b latency was laterin older adults relative to younger adults underhigh WML. Older adults showed larger con-tralateral activation and lower ipsilateral inhibi-tion of motor areas than younger adults. TheN1 in occipital visual cortex was larger in olderthan in younger adults. FB facilitated perfor-mance under high WML in both groups that
2010, 3 (1) Kognitive Neurophysiologie des Menschen 41

W. Skrandies — Abstracts of the 18th German EEG/EP Mapping Meeting
were accompanied by different neuronal pat-terns. In younger adults FB led to shorterP3b latency, while no changes in motor acti-vation was found. In older adults FB did notchange P3b latency and delayed motor acti-vation. FB positivity was later in older com-pared to younger adults. The results suggestthat both age group benefit from FB in termsof performance improvement was achieved indifferent ways. Younger adults seem to imple-ment FB better for chunking information. Inthis way the categorization process was facil-itated and resulted in earlier P3b. In contrast,FB in older adults provokes the adoption of anaccuracy-oriented strategy when an impairedcategorization process is compensated for byenhanced attention, facilitation the early en-coding process and delaying of motor activa-tion. In sum, FB appears to facilitate a reor-ganization of WM in younger adults and to en-hance controlled processing in older adults.
Measuring compensation and its plasticityacross the lifespan.J. Zöllig, Department of Psychology, Univer-sity of Zurich, Zurich, Switzerland.Two important aspects have to be inte-grated when studying compensation: (1)the knowledge about specific compensationalmechanisms and (2) the consideration oflifelong changes in these mechanisms, i.e.,the plasticity of compensation. Accordingly,the present talk addresses the questions(a) which neural networks are supportingsuccessful cognitive performance acrossdevelopment, (b) what are the associatedcompensational mechanisms and (c) arethese compensational mechanisms plasticacross the lifespan. To answer these ques-tions, we suggest to integrate behavioural and
neuroimaging methods and present specificmethods and their advantages and disadvan-tages. The relevance of this integration willbe illustrated by presenting data from our labusing ERPs and (s)LORETA to study com-pensational processes across the lifespan ina higher order cognitive function, i.e., delayedintentional behaviour. A higher activation inold adults or adolescents in successful tri-als compared to young adults is consideredcompensatory as specialized cortical regionsare selectively recruited in response to taskdemands. The findings from our lab suggestthat whereas the performance outcome mightbe the same in different age groups, un-derlying processes and activations might befundamentally different. The talk will concludewith a discussion about specific implicationsof this integrative approach when studyingthe adaptive potentials and limits of humancognition.
Age-related differences in a driving-likedual task situation.N. Wild-Wall, M. Hahn & M. Falkenstein, Leib-niz Research Centre for Working Environmentand Human Factors, Dortmund, Germany.Research revealed age-related deficits invisuo-motor dual tasks. Car driving can betaken as a related practical example. Bymeans of behavioural- and EEG-data weinvestigated central processes in young andolder participants during a driving-like dualtask. Participants performed a primary track-ing task and a secondary visually cued atten-tion task. In the visual attention task underdual task condition, the older vs. young par-ticipants showed generally higher responsetimes, however, comparable error rates. Inaddition, they improved their behavioural per-
42 Human Cognitive Neurophysiology 2010, 3 (1)

W. Skrandies — Abstracts of the 18th German EEG/EP Mapping Meeting
formance more than the younger participants.A higher CNV amplitude of the old vs. younggroup between cue and stimulus suggeststhat the older invested more ressources tothe dual task in this interval. By means ofsLORETA the age-correlated amplitude dif-ference can be explained by higher activationof the older vs. young participants in rightparietal, bilateral frontal and postcentral areasas well as in the precuneus. The trackingerror of the older vs. young participants wasgenerally increased, and relatively more in-creased after relevant targets in the attentiontask. In contrast, their tracking error relativeto their individual tracking performance waseven lower compared to the young group inthe cue-stimulus interval. Possibly, the olderuse the invested ressources in this intervalnot only to prepare for the following stimulus,but also, to meet the task requirements of thetracking task in the difficult dual task situa-tion. In sum, older participants were able toimprove their performance in some aspectsof the dual task by means of practice andeffort. Nevertheless, the dual task situationcan lead to a performance decline in thetracking task especially after relevant targetsin the secondary attention task. The resultsbear practical relevance e.g. for the design ofinformation and assistence systems for olderdrivers.
Simultaneous wavelet analysis of EEG-channels.A. Klein (1), T. Sauer (1) & W. Skrandies(2), (1) Lehrstuhl für Numerische Mathematik,Heinrich-Buff-Ring 44, D-35392 Gießen; (2)Physiologisches Institut Aulweg 129, D-35392Gießen, Germany.One-dimensional wavelet analysis allows
for the processing of single channels only.Hence, any suspected connection betweenchannels has to be analysed either before orafter the transform for every pair of channels,making the analysis very cumbersome attimes. Matrix-wavelets, however, provide anovel means for the analysis of vector-valuedsignals, i.e. of several channels at once, andcan even be parametrised so as to take intoconsideration the relations between chan-nels. While the computations employed inthe method are very intricate, they are hiddenfrom the user, and in the standard case allinformation contained in the data is preservedduring the transform. This allows any ab-straction from the data to be pushed to laterstages in the analysis, making the analysismore robust to unobvious errors, for exampleincorrect indexing. We present first resultsillustrating how the choice of parameterscan influence the reduction of dimensionalitybased on PCA.
Sparseness in space meets sparseness intime: EEG sychronization is associatedwith fMRI resting state networks.T. Koenig, K. Jann, M. Kottlow, W. Strik, C.Boesch & T. Dierks, Abteilung für Psychia-trische Neurophysiologie, Universitätsklinikfür Psychiatrie und Abteilung für klinis-che Forschung / AMSM, Universität Bern,Schweiz.FMRI resting state data display several sets ofco-activated brain regions (so called RestingState Networks; RSNs). EEG resting statedata show a limited set of highly synchronizedscalp field patterns, suggesting transientphase-locking of distributed cortical patches(Koenig, 2005). We investigated whether theknown fMRI RSNs coincide with EEG-fMRI
2010, 3 (1) Kognitive Neurophysiologie des Menschen 43

W. Skrandies — Abstracts of the 18th German EEG/EP Mapping Meeting
BOLD correlates of specific patterns of phase-synchronous EEG. Combined resting-state96-channel EEG and 3T-fMRI was recorded in14 healthy young subjects. fMRI RSNs wereidentified by an ICA. The EEG was analyzedusing a) a global frequency domain measureof synchronization (GFS) and b) wavelet anal-yses combined with spatial clustering to iden-tify transiently oscillating synchronous EEGstates (microstates). GFS and microstatesdynamics were used as regressors on thefMRI signals to identify their BOLD correlates.Those BOLD correlates were then comparedto the previously identifed RSNs. GFS BOLDcorrelates in lower alpha (8.5-10.5Hz) coin-cided with the Dorsal Attention RSN, whileGFS BOLD correlates of the upper alphaband resembled the Default Mode Network.The dynamics of different microstate classeswere also associated with the Default ModeNetwork, a network associated with frontalattention and self-reference, and others. Con-clusions: Specific patterns of common-phaseEEG oscillations are associated with specificpatterns of fMRI co-activation during rest.This supports the hypothesis that the differ-ent modules of widespread neuro-cognitivenetworks observed during rest (and obvi-ously also during task execution) are coupledthrough a transient phase-locking of theiroscillations.With support of the Swiss National Science Foun-
dation grant 320000-108321/1.
EEG source localization differences be-tween adult and pediatric realistic headmodels.M. Wagner, M. Fuchs & J. Kastner, Com-pumedics Neuroscan, Hamburg, Germany.Even if individual MRI data and digitized elec-
trode positions are unavailable, realistic headmodels yield more accurate source localiza-tions than spherical shells [1]. In this contribu-tion, source localization differences betweenadult realistic head models derived from theICBM-152 geometry [2] and pediatric headmodels derived from the MNI-175 geometry[3] are elaborated. The respective roles ofhead shape and skull conductivity are quan-tified.[1] M. Fuchs, M. Wagner, J. Kastner. 2001. Boundary el-ement method volume conductor models for EEG sourcereconstruction, Clinical Neurophysiology 112:1400-1407.
[2] J. Mazziotta et al. 2001. A probabilistic atlas andreference system for the human brain. PhilosophicalTransactions of the Royal Society B: Biological Sciences356:1293-1322.
[3] http://www.bic.mni.mcgill.ca/cgi/icbm_view
Pinprick-evoked potentials: Correlate ofpositive symptoms in nociception.
U. Baumgärtner, G. Iannetti, W. Magerl, I.Tracey & R.-D. Treede, Adressen: Lehrstuhlfür Neurophysiologie, Zentrum für Biomedizinund Medizintechnik Mannheim (CBTM), Med.Fak. Mannheim der Universität Heidelberg,Mannheim, Deutschland; Institute of Physiol-ogy, Anatomy and Genetics, Oxford Univer-sity, Oxford, United Kingdom.
To date, nociceptive pathways can be func-tionally evaluated using laser-evoked poten-tials. However, these signals are sensitiveto detect lesions by means of neurophysio-logic deficits (reduced or delayed amplitudes),whereas an objective measure for plus-symptoms like mechanical hyperalgesia ismissing so far. Here we describe a noveltechnique, pinprick-evoked potentials (PEPs),which may close this clinically important gap.PEPs were elicited by mechanical stimulationusing a flat-tip probe (diameter 0.25 mm,
44 Human Cognitive Neurophysiology 2010, 3 (1)

W. Skrandies — Abstracts of the 18th German EEG/EP Mapping Meeting
force 128 mN), which activates A-delta no-ciceptors, and is frequently used to clinicallyassess mechanical pain sensitivity. A triggeroutput enabled us to average the responsestime-locked to the onset of the mechanicalstimulation. PEPs were recorded in 10 sub-jects during stimulation of the right and lefthand dorsum before and after intradermal in-jection of capsaicin into the right hand, whichexperimentally induces mechanical hyperal-gesia in the skin surrounding the injectionsite. PEPs in response to stimulation of nor-mal skin were characterised by large biphasicnegative-positive (NP) complex, maximal atthe vertex, with the N and P waves havingapproximate latencies of 100 ms and 220ms, and amplitudes of 5 V and 12 V, respec-tively. All subjects developed mechanicalhyperalgesia following capsaicin injection.Stimulation of the hyperalgesic skin resultedin a significant N amplitude increase (+63.4%,p=0.02, one-sample t-test). The increaseof the P wave was not significant (+15.3%,p=0.10). In one patient with a high spinallesion and reduced unilateral pain sensitivitybelow the lesion, standard SEP were normaland symmetric, whereas PEP amplitudes aswell as laser-evoked potentials were reducedon the symptomatic side. These results in-dicate that PEPs (1) primarily reflect corticalactivities triggered by somatosensory inputtransmitted in A-delta primary sensory affer-ents and spinothalamic projection neurons,and (2) represent a useful tool to exploreexperimentally-induced secondary mechani-cal hyperalgesia.
Contact cool and heat-evoked potentials -CEPs and CHEPs - are mediated by distinctA-fiber classes.
W. Greffrath, D. Pfau, W. Tiede, U. Baumgärt-ner & R.-D. Treede, Lehrstuhl für Neurophys-iologie, Zentrum für Biomedizin und Medizin-technik Mannheim (CBTM), Universitätsmedi-zin Mannheim, Ruprecht-Karls-UniversitätHeidelberg, 68167 Mannheim, Deutschland.
Perception of non-painful cold stimuli and ofpricking pain plays an important role in quan-titative sensory testing (QST) as parametersfor A-delta-fiber function. Since QST de-pends on active participation of the patient,objective tests for A-delta-fiber function arealso needed. Whereas laser-evoked poten-tials (LEPs) and contact-heat evoked poten-tials CHEPs have been validated, no suchtest exists for the thermoreceptive pathwaysfor cooling. Evoked potentials elicited by acontact-thermode stimulator (CHEPS, MedocLtd.) were recorded from scalp electrodes.Cool (by -3 deg C) and heat stimuli (by +16deg C) were directed bilaterally to the handdorsum of 12 healthy volunteers from a base-line temperature of 35 deg C in 16 blocksof 20 stimuli each (ISI 8-10s). Subjectiveheat pain/coolness of each stimulus was sub-jectively rated using a numeric rating scale.Mean heat pain ratings were 23.1 (SD 6.3),cooling was not perceived as painful, meancoolness ratings were 3.4 (SD 3.0). Precededby a small negativity (N1), the evoked po-tential for both stimuli consisted of a vertexnegativity (N2) followed by a positivity (P2).Peak amplitudes did not differ between cooland heat evoked potentials (15.99 (SD 2.06)V versus 19.17 (SD 2.27 V; p>0.05). La-tencies were shorter with cooling than heat-ing (N2 323 vs 397ms, t-test; p<0.01). Heatpain ratings steeply decreased initially (tau1about 15 s) and leveled out thereafter (tau2
2010, 3 (1) Kognitive Neurophysiologie des Menschen 45

W. Skrandies — Abstracts of the 18th German EEG/EP Mapping Meeting
about 88 s), cool ratings dropped very slowly(tau>2000s). Local desensitization with 10%capsaicin creme for >24 h completely abol-ished both, heat pain and CHEPs but did notreduce subjective coolness and CEPs. Thesedata indicate that both stimuli elicit similar ver-tex potentials via activation of distinct A-deltafibers. Peripheral adaptation to cool stimuliis less pronounced than to heat. CombiningCEPs with CHEPs could be used as objectivemethod to examine somatosensory pathwaysin more detail.
Supported by DFG Tr236/11-3 and 13-3.
Mapping the cortical correlates of acous-tic lateralization with Near-Infrared Spec-troscopy.
J. Zeller (1), W. Harnisch (2), M. Plichta(1), E. Hartwig (2) & A. Fallgatter (1), (1)Psychophysiologie und funktionelle Bildge-bung, Klinik für Psychiatrie, Psychosomatikund Psychotherapie der Julius-MaximiliansUniversität Würzburg (2) Klinik u. Polik-linik für Hals-, Nasen- u. Ohrenkrankheiten,plastische u. ästhetische Operationen derJulius-Maximilians Universität Würzburg, Ger-many.
Interaural time differences (ITD) are ma-jor cues for the spatial perception of soundsources. In normal hearing subjects, binauralpresentation of acoustic stimuli leads to a shiftof sound perception to either left or right side,depending on the degree of time difference(lateralization). There is growing evidencethat the processing of acoustical stimuli isorganized in a cortical dual pathway systemin which certain areas are responsible for theprocessing of qualitative stimulus propertieswhereas other areas are involved in the pro-cessing of spatial information. During the
localisation of sounds fMRI studies show ac-tivation of the inferior parietal cortex and thesuperior frontal sulcus. However, an objectivemeasure of spatial hearing ability is neededfor patients that cannot be easily assessed inthe scanner (i.e. paediatric patients, cochlearimplant users). The aim of the present studywas to investigate cortical correlates of later-alization in a group of healthy controls withnear-infrared spectroscopy (NIRS) that of-fers considerable advantages in setting foracoustical paradigms (no scanner noise, noartefacts due to metal implants). 50 adultnormal hearing subjects completed a soundlateralization task, where subjects indicatedthe perceived direction (left, right, frontal) ofan acoustic stimulus (100µs clicks or 830msclicktrains, 50 dB SL) presented via insertableheadphones. Simultaneously changes in cor-tical oxygenation were measured using 2x22channel NIRS over parietal and frontal areas.Comparing stimuli with large ITD (750 µs) tosuch with shorter ITD (100 µs) we observedan activation of the left inferior parietal cortex.Thus NIRS seems to be a suitable methodto measure the cortical correlates of acousticlateralization.
Early electrophysiological effects duringStroop tasks – First results.W. Skrandies & S. Iranpour, PhysiologicalInstitute, Justus-Liebig University, D-35392Giessen, Germany.According to brain imaging studies, non-compatible Stroop stimuli activate the anteriorcingulum of the human brain. In electrophys-iological experiments effects on long-latencycomponents (400-800 ms) are stressed. Weinvestigated 16 adults who processed com-patible and non-compatible color words which
46 Human Cognitive Neurophysiology 2010, 3 (1)

W. Skrandies — Abstracts of the 18th German EEG/EP Mapping Meeting
were presented on a monitor. Presentationwas randomized, and stimuli occurred in thecenter, or in the left or right visual field. EEGwas EEG was recorded from 30 channelslocated between the inion and 5Between 80and 140 ms latency there was a significantinteraction between compatibility and visualfield location (F(2,30) = 3.89, p < 0.05), andbetween 150 and 250 ms component la-tency was higher with compatible than withnon-compatible stimuli (F(1,15) = 4.55, p <0.05). The third component occurred between250 and 350 ms latency, but it showed nosignificant effects. Spatial PCA resulted infour components that explained 78.6% of thevariance. Factor scores were employed indiscriminant analyses in order to identifiedexperimental conditions. For ERPs elicitedby central stimuli, at 110 ms latency 75Ourresults show how the compatibility of Stroopstimuli is reflected in field strength and topog-raphy as early as at about 100 ms latency. Inaddition to late components reported in theliterature there is rapid information processingin primary visual areas during Stroop tasks.
EEG spectral power before and duringhypnosis reflects hypnotizability.C. Andreou (1), D. Lehmann (2), P. Jon-sson (3), P.L. Faber (2), D.B. Terhune (3)& E. Cardena (3), (1) 1st Psychiatric De-partment, Aristotle University of Thessaloniki,Greece; (2) The KEY Institute for Brain-MindResearch, University Hospital of Psychiatry,Zurich, Switzerland; (3) Dept. of Psychology,University of Lund, Sweden.Healthy people volunteering for hypnosis dif-fer in hypnotizability. EEG data that mightreflect this predisposition before baselineand during hypnosis were controversial.
Our multichannel-EEG study intends to clar-ify the controversies. 60-channel EEG wasrecorded from eleven high (’highs’) and elevenlow (’lows’) hypnotizable healthy volunteerscomfortably seated with eyes closed. EEGrecordings before, immediately after induc-tion of hypnosis, and in deep hypnosis werespectral analyzed (relative power versus av-erage reference) in eight frequency bands.Subjects self-rated their hypnotic depth. T-tests and repeated-measures ANOVAs wereapplied for exploratory statistics. Before hyp-nosis, highs and lows showed no significantdifference in self-rated hypnosis depth, butfor deep hypnosis, highs rated depth signif-icantly higher than lows. Before hypnosis,highs had lower power in the theta and al-pha1 frequency bands, but higher power inbeta3. Single-channel analyses revealedno differences between hemispheres. Thechanges of power spectra over the threetime points exhibited the following differencesbetween groups: Highs mainly showed anincrease in theta power (F=4.48, p=0.03),while lows showed a decrease in alpha1(8.5-10 Hz) [F=8.64, p=0.01], an increase inalpha2 (10.5-12 Hz) [F=7.13, p=0.005], andfluctuations in gamma frequency (35-45 Hz)power [F=3.89, p=0.04]. Our results agreewith earlier reported changes in theta andalpha EEG frequency bands that distinguishlow- and high-hypnotizable subjects beforeand during hypnosis, and that suggest higherarousal in highs before hypnosis.Supported partly by Bial Foundation
Are short reaction times related to frontalmidline theta?B. Artner, E. Weber & M. Doppelmayr, Deptof Physiological Psychology, University of
2010, 3 (1) Kognitive Neurophysiologie des Menschen 47

W. Skrandies — Abstracts of the 18th German EEG/EP Mapping Meeting
Salzburg, Hellbrunnerstr 34., 5020 Salzburg,Austria.
Inconsistent results have been reported cor-relating EEG parameters with reaction time(RT) (Oken et al. 2006). Due to the factthat attention is a prerequisite for fast reac-tions and is associated with the frontal mid-line theta, we investigated this relationship offrontal theta power and RT. 35 healthy sub-jects (11m, 24 f) performed two visual sim-ple RT-tasks, with either a fixed (4000 ms) ora variable inter-stimulus interval (ISI) (3500 -5750 ms). Theta frequency was adjusted in-dividually from 6 - 4 Hz below the individualalpha peak frequency (Klimesch 1999). Theprestimulus interval (-750 to 0 msec) was di-vided in 3 x 250 msec segments (t1, t2, andt3). Data were analyzed for 5 midline positionsonly. RTs below 125 ms or exceeding 1000ms were excluded, and short/long RTs weredivided according to the individual median RT.A four factorial ANOVA including the factorsPERFORMANCE (fast/slow), TASK (fixed vsvarying ISI), TIME (t1, t2, t3), and LOCA-TION (Fz, FCz, Cz, Pz, and Oz) was con-ducted. Besides significant main effects forPERFORMANCE, TIME and LOCATION aswell as several 2-factorial interactions, themost interesting finding was a significant in-teraction of TASK x PERFORMANCE x TIME.While for varying ISI thetapower remained un-changed over time and performance, in thefixed ISI condition thetapower increased fromt1 to t3. Interestingly this increase was morepronounced for fast RTs. These results canbe interpreted in a way that, while with vary-ing ISI the subjects are unable to prepare cor-rectly for a fast response, this is possible for afixed ISI. In addition, if in the fixed ISI condi-
tion, prestimulus thetapower is increased, fastresponses are more likely as compared to tri-als with lower power.Klimesch W., (1999) Brain Res. Rev. 29, 169-195.
Oken, B.S., Salinsky, M.C., & Elsas, S.M., (2006)
Clin.Neurophysiol. 117, 1885-1901
EEG power spectra, LORETA areas andself-rated headache pain.P. L. Faber (1), S. Tei (1,2), C. Chen (3), P.Hsiao (3) & D. Lehmann (1), (1) The KEY Insti-tute for Brain-Mind Research, University Hos-pital of Psychiatry, Zurich; (2) Department ofStress Science and Psychosomatic Medicine,Graduate School of Medicine, The Universityof Tokyo, Japan; (3) Dept. of Neurology, TaipeiVeterans General Hospital, Taiwan.How is the subjective intensity of headachereflected in EEG power spectra, and wherein the brain is the origin? In 16 migrainepatients (without aura, 6 medicated; meanage 38.3 years, SD=7.7; 12 women), werecorded 19-channel EEG during initial rest-ing (4 minutes) on an attack-free day. Patientsvisual-analog self-rated headache intensity.On average/patient, 65.6 s artifact-free EEGwas available. For each of the eight inde-pendent EEG frequency bands, means ofspectral power across channels (absoluteand relative), and LORETA functional im-ages (absolute and relative voxel strength)were computed, and correlated with sub-jective headache intensity. The correlationswere negative (p<0.1) with absolute power inthe delta, alpha2, beta1 and 3, and gammabands, but positive (p<0.1) with relative powerin delta and beta2. Absolute LORETA voxelstrength (corrected for multiple testing p<0.1)correlated negatively in all bands but delta;relative voxel strength correlated positively
48 Human Cognitive Neurophysiology 2010, 3 (1)

W. Skrandies — Abstracts of the 18th German EEG/EP Mapping Meeting
with delta through alpha1. Negative cor-relations with absolute LORETA strengthconcerned only left anterior voxels in thetaand alpha1, only left-hemispheric in beta1and beta2, predominantly left in alpha1 andbeta3, but more right in gamma; all positivecorrelations with relative strength were rightanterior (delta, theta and alpha1). Medicatedand unmedicated patients showed agreeingtendencies. Thus, with increasing headacheintensity, EEG absolute power decreased,localizing in the left hemisphere (some fre-quency bands predominantly anterior), whilerelative strength increased exclusively rightanterior. The results suggest that increas-ing headache intensity is associated withincreasing left- hemispheric anterior-weightedactivation.Supported in part by Bial Foundation Grant 44-06
Time course of mental calculation pro-cesses: a combined fMRI and MEG study.T. Fehr (1,2), H. Hinrichs (3), C. Code (4) & M.Herrmann (1,2), (1) Dept. of Neuropsychol-ogy/ Behavioral Neurobiology, Center for Cog-nitive Sciences, University of Bremen, Bre-men, Germany; Center for Advanced Imaging,Bremen/Magdeburg, Germany; (3) Dept. ofNeurology, University of Magdeburg, Magde-burg, Germany; (4) School of Psychology,University of Exeter, Exeter, UK.The issue of how arithmetic operations arerepresented in the brain has been addressedin numerous studies. Lesion studies suggestthat a network of different brain areas (e.g.the left and/or right parietal lobe, frontal cor-tex, and basal ganglia) is involved in mentalcalculation processing. Neuroimaging stud-ies have reported inferior parietal and lateralfrontal activations during mental arithmetic
using tasks of different complexities and us-ing different operators. There is, however,scarce information about the time course ofsequential activation of the different brainregions involved in mental arithmetic. Thepresent study investigated fMRI-BOLD andevent-related MEG activity during addition,subtraction, multiplication and division per-formance in tasks of differing in complexity.Tasks were presented both auditory and visu-ally in two different runs within one session.Modality of stimulus presentation resulted insubstantially different BOLD activation pat-terns. Data showed activation patterns inprominently right frontal, parietal and sub-cortical regions when contrasting complexand simple calculation tasks. Preliminaryfindings demonstrate different task-relatedspatio-temporal characteristics of dipole waveforms as revealed by seeded dipoles basedon fMRI activation foci. However, as largevariances in single trial based data analysessuggest individual different time courses ofactivity during complex mental calculation, itis suggested that MEG data, obtained duringcomplex mental processing, should betterbe analysed on a non-averaging-based sin-gle trial level considering potential individualdifferences in task-related oscillatory brainactivity and variations in timing of mentalprocessing of cognitive sub-components.
Gender related differences in visual pro-cessing.N. Jausovec & K. Jausovec, University ofMaribor, Faculty of Arts, 2000 Maribor, Slove-nia.The aim of the present study was to investi-gate gender related differences in brain ac-tivity for tasks of verbal and figural content
2010, 3 (1) Kognitive Neurophysiologie des Menschen 49

W. Skrandies — Abstracts of the 18th German EEG/EP Mapping Meeting
presented in the visual and auditory modal-ity. Thirty male and 30 female respondentssolved 4 tasks while their electroencephalo-gram (EEG) was recorded. Also recordedwas the percentage of oxygen saturation ofhemoglobin (%StO2 ) in the respondents?frontal brain areas with near-infrared spec-troscopy (NIRS). The main findings of thestudy can be summarized as follows: (1) Mostpronounced differences between males andfemales were observed for the factor modal-ity - visual/auditory. (2) Gender related differ-ences in neuroelectric brain responses couldbe observed during the solution of auditoryand visual tasks, however on the behaviorallevel only for the visual tasks did females dis-play shorter reaction times than males. TheERP amplitudes of the early evoked gammaresponse, P1, and P3 were higher in femalesthan males, whereas the N4 amplitude washigher in males than females. The differenceswere more noticeable in the visual modality.The NIRS showed a more bilateral involve-ment of the frontal brain areas in females ascompared with a more left hemispheric frontalactivity in males. In the task conditions an in-crease in right hemispheric activity in femaleswas observed, however this increase was lesspronounced in the visual than the auditory do-main, indicating a more lateralized processingof visual stimuli in females. Taken all togetherthe results suggest that the females’ visualevent-categorization process is more efficientthan in males.
Event-related alpha oscillations related toperception, sensorimotor processing andresponse inhibition in elderly.Ch. Schmiedt-Fehr & C. Basar-Eroglu, Insti-tute of Psychology and Cognition Research,
University of Bremen, Germany.
A major challenge for developmental cog-nitive neuroscience is to understand howchanges in cognitive functions related toaging are associated with changes in theneuronal information processing architecture.Previous studies on EEG event-related brainoscillations suggest functional changes inalpha-bands with age during sensory andmemory tasks. The topographical distributionof both single trial lower and upper alphamagnitude and the corresponding phasecoherence is documented to be altered inelderly persons. Thus, alpha oscillations,not only associated with sensory, but alsowith sensorimotor functions may be alteredwith age. Thereby, compensatory mecha-nisms, possibly reflected in increased frontalalpha synchronization, may be of profoundrelevance. The present study investigatesage-related differences in the modulation ofalpha oscillatory activity related to sensoryand sensorimotor functions, including re-sponse preparation, execution, and inhibition.EEG was recorded while ten young and tenelderly persons performed a visual evokedpotential (VEP), a cued stimulus response(S-R) and a cued visual Go/NoGo task. Earlyevent-related synchronization (ERS, 0-250ms) and late event-related desynchroniza-tion (ERD, 200-600 ms after stimulus onset)of single trial lower and upper alpha wasanalysed using poststimulus amplitude en-hancement and inter-trial phase coherencemeasures. The results show comparablemodulations of lower (8-10 Hz) and upper(10-15 Hz) alpha activity in young and elderlyrelated to the sensory and different forms ofmotor response processing. Specifically, less
50 Human Cognitive Neurophysiology 2010, 3 (1)

W. Skrandies — Abstracts of the 18th German EEG/EP Mapping Meeting
upper alpha ERD at central locations andreduced early lower alpha ERS character-ized response inhibition processing in bothage-groups. We discuss the counter-intuitive,hypothesis that reduced efficacy of somebasal neural inhibitory mechanisms could en-able older observers to perform better than orcomparable to younger persons on tasks us-ing large, high contrast checker board stimuli.We conclude that observation of alterationsin alpha oscillatory networks with age maydepend on the stimuli applied.
Error-related components and impulsivityreflected by trade-off between speed andaccuracy.H. Shinoda (1) & K. Suzuki (2), (1) Depart-ment of Clinical Psychology, Faculty of Psy-chology, Rissho University, (2) Department ofPsychology, Graduate School of Psychology,Rissho University, Tokyo, Japan.We examined a neurophysiological aspect ofthe impulsivity as defined by performance.First, we selected 71 participants from agroup of undergraduate and graduate stu-dents. They performed an arrow versionof flanker task and were classified into 3subgroups by determining RT and the rateof incorrect response: the impulsive group(+1 SD above average or higher in incorrectresponse rate), the average group (+1 SDbelow average or shorter in RT and lower inincorrect response rate) and the remaininggroup. Next, the flanker tasks were adminis-tered to 7 participants from the impulsive (6male and 1 female) and the average group (5male and 2 female) during EEG recordings.We consistently confirmed that individuals inthe impulsive group had shorter correct RTand higher incorrect response rate for incom-
patible stimuli compared to individuals in theaverage group. Furthermore, double-errorrate was significantly higher in the impulsivegroup than in the average group. We foundtwo clear GFP peaks following response inboth groups, and these components wereidentified as ERN/Ne and Pe. Although thevalue of GFP showed no difference in bothgroups, the amplitude of ERN/Ne at Cz wassignificantly lower in the impulsive group ascompared to the average group. In scalp fieldtopography, we noticed that the negative cen-troids at ERN/Ne peak was central area in theaverage group while it was more anterior areain the impulsive group. Psychophysiologicaldata showed lower ERN/Ne activity over cen-tral area in the impulsive group. These resultssuggest that the inability to adjust behaviorunderlying impulsivity is associated with theERN/Ne activity.
Training in breath counting is reflected byEEG power and LORETA functional im-ages.S. Tei (1,2), C. Chen (3), P.L. Faber (1), P.Hsiao (3) & D. Lehmann (1), (1) The KEYInstitute for Brain-Mind Research, UniversityHospital of Psychiatry, Zurich, Switzerland, (2)Department of Stress Science and Psychoso-matic Medicine, Graduate School of Medicine,The University of Tokyo, Japan, (3) Dept. ofNeurology, Taipei Veterans General Hospital,Taiwan.Does breath count training alter brain electricactivity? Breath counting is a basic exercisefor meditation. Since meditation has beensuggested as migraine treatment, we wantedto determine whether breath count trainingaffects brain activity assessed with EEG.On attack free days, 19-channel EEG were
2010, 3 (1) Kognitive Neurophysiologie des Menschen 51

W. Skrandies — Abstracts of the 18th German EEG/EP Mapping Meeting
recorded from 16 migraine patients (meanage 38.3 years, SD=7.72; 12 women) duringinitial resting, breath counting (repeatedlycount one to ten for 5 minutes) and finalresting, all with eyes closed. Seven of thesepatients were assigned to do one month ofbreath count training; thereafter, all EEGrecordings were repeated. EEG was spectralanalyzed versus average reference; mean rel-ative power across channels was computedin the eight independent frequency bands.EEG-LORETA funtional imaging was used tolocalize changes observed in the head sur-face EEG recordings. ANOVA’s (2 sessions x2 conditions x 2 groups) for the 8 frequencybands revealed no main effects, but an inter-action (p=0.058) in the theta frequency band.In this band, post-hoc tests showed, duringthe second session, a significant power in-crease from initial to final resting only in thetrained group. LORETA functional tomogra-phy revealed that the observed differencesconsisted in significantly (corrected for mul-tiple testing) increased voxel strength in leftparietal areas. In sum, state-dependent learn-ing effects were observed in the EEG afterone-month training of breath counting: theseeffects occurred not in the resting state, but inthe state immedialy after having completed abreath count exercise.Partly supported by Bial Foundation 44-06
Neurofeedback for children with attentiondeficit / hyperactivity disorder (ADHD):Clinical and neurophysiological results ofa randomised controlled trial.P. Studer (1), H. Gevensleben (2), S. Wan-gler (1), A. Rothenberger (2), G.H. Moll (1)& H. Heinrich (1,3), (1) Kinder- u. Jugend-abteilung für Psychische Gesundheit, Univer-
sitätsklinikum Erlangen; (2) Kinder- u. Ju-gendpsychiatrie, Universitätsklinikum Göttin-gen; (3) Heckscher-Klinikum, München, Ger-many.
Neurofeedback is a neuro-behavioural,computer-assisted training which could be-come an important treatment module forchildren with attention deficit/hyperactivitydisorder (ADHD). In a multisite randomisedcontrolled study, we evaluated clinical andneurophysiological effects of a neurofeedbacktraining compared to a control training. 102children with ADHD (age: 8 to 12 years) par-ticipated in the study. They either performeda neurofeedback training or a computerisedattention skills training (randomised groupassignment). The training programs com-prised 36 sessions within two blocks of about4 weeks each. The neurofeedback train-ing consisted of both a theta/beta trainingand a slow cortical potentials (SCP) training.Pre-training, intermediate and post-trainingassessment comprised several behaviourrating scales (e.g. the German ADHD rat-ing scale, FBB-HKS) and neurophysiologicalmeasurements (EEG, event-related poten-tials). For both parent and teacher ratings,the neurofeedback training was superior tothe control training. The effect size was .60for the FBB-HKS total score (parent ratings,primary outcome measure). In the restingEEG the neurofeedback group showed areduced theta activity after the end of train-ing. In addition, EEG measures recorded atbaseline as well as changes of EEG param-eters from pre to post training were relatedto the clinical outcome. Furthermore, SCPtraining was associated with an increase ofthe contingent negative variation in a cued
52 Human Cognitive Neurophysiology 2010, 3 (1)

W. Skrandies — Abstracts of the 18th German EEG/EP Mapping Meeting
attention test. Contrary to our expectations noincrease of the P300 amplitude was found forthe theta/beta training group. Neurofeedbackcan be considered as a clinically effectivemethod for the treatment of children withADHD. The neurophysiological effects couldreflect correlates of a successful training. Fu-ture studies should examine how to optimisea neurofeedback training and how to integrateit into a multimodal treatment program forchildren with ADHD.Gevensleben H, Holl B, Albrecht B, Vogel C, Schlamp D,Kratz O, Studer P, Rothenberger A, Moll GH, HeinrichH (2009): Is neurofeedback an efficacious treatment forADHD? A randomised controlled clinical trial. J Child Psy-chol Psychiatry, 50: 780-789.
Funded by the German Research Foundation
(HE4536/2, MO 726/2, RO 698/4)
Effects of SMR and sham neurofeedbackon EEG amplitude.M. Doppelmayr, E. Weber & K. Hödlmoser,Department of Physiological Psychology, Uni-versity of Salzburg, Salzburg, Austria.During the last few years several studies haveshown that humans are able to modify theirown brain rhythms by the means of neuro-feedback (NF). Besides the meanwhile estab-lished use of NF in therapeutical settings asfor the treatment of ADD/ADHD or epilepsy,the effects of NF in healthy subjects are lessclear. Especially for healthy subjects, there isa lack of empirical data focusing on the ef-fects of NF on the EEG. Although there areseveral reports indicating that subjects areable to learn to increase the amplitude of thesensorimotor rhythm (SMR; 12-15 Hz), thereare almost no studies describing the con-comitant effects on a resting EEG precedingand/or following NFT. In the reported study 12healthy subjects received visual feedback of
the SMR while the 8 subjects of the controlgroup received feedback from several vary-ing frequency bands (pseudo-neurofeedbackPNF). Both groups performed 25 NF-trainingsessions (5 weeks with 5 trainings) and wereinstructed to increase the power as depictedby an in- or decreasing circle. EEG wasrecorded at C3 and C4 during the training aswell as in resting conditions preceding and fol-lowing each NF session. The results indicatedthat, during training, SMR amplitude signif-icantly increased in the experimental group,whereas it remained unchanged in the PNF-training. However, this increase in SMR am-plitude was present only during the training.Interestingly there was also a strong tendencytowards increased SMR amplitudes during theeyes open condition.
Project ADHD & EEG-Neurofeedback Ther-apy (PANther).M. M. Lansbergen (1), M. van Dongen-Boomsma (1,2), D. Slaats-Willemse (1,2) &J. K. Buitelaar (1,2), (1) Radboud UniversityNijmegen Medical Centre, Donders Institutefor Brain, Cognition and Behaviour, Depart-ment of Psychiatry, Nijmegen, The Nether-lands; (2) Karakter University Centre for Childand Adolescent Psychiatry, Nijmegen, TheNetherlands.Attention Deficit Hyperactivity Disorder(ADHD) affects approximately 4% to 12% ofall school-age children. Primary treatment forADHD is medication, although 30% of childrenfail to respond to psychostimulants. Findingsfrom previous studies suggest that electroen-cephalography (EEG)-neurofeedback trainingis a promising alternative treatment for ADHD.However, few studies evaluated the effectsof neurofeedback treatment in comparison
2010, 3 (1) Kognitive Neurophysiologie des Menschen 53

W. Skrandies — Abstracts of the 18th German EEG/EP Mapping Meeting
with a control treatment that corrects forexpectancy effects and other unspecific fac-tors. This project involves a double-blind,randomized, placebo controlled study in 120children with ADHD (8-15 years) with as mainobjective to examine the effects of EEG neu-rofeedback in ADHD on clinical symptomsof ADHD and cognitive functions, controllingfor unspecific treatment effects. A secondaryobjective is to examine the underlying mecha-nisms of EEG neurofeedback. ADHD childrenwill receive 30 sessions of personalized EEGneurofeedback or sham feed back (i.e., feed-back based on a simulation signal). Priorto and after treatment, rating scales will befilled out, neuropsychological measures willbe administered, and EEG and (f)MRI mea-surements will be performed. So far, 15children participated in this study. Preliminaryfindings of pre- and post assessments will bepresented. Since it is a double-blind study,the preliminary results will be presented with-out differentiating between groups. Our studywith a double-blind, randomized, placebo con-trolled design will make a novel contributionto the literature about the effects of neuro-feedback in ADHD. Moreover, the acquisitionof MRI and EEG data before as well as afterneurofeedback/sham training may give moreinsight into the mechanisms underlying EEGneurofeedback.
ADHD Neurofeedback Research NetworkSymposium: Preliminary tomographicneurofeedback results from children withADHD.M. Liechti (1,2), S. Maurizio (1), H. Heinrich(3,4), G. Thalmann (1), L. Meier (1), Y. Schwit-ter (1), M. Hartmann (1), S. Walitza (1), H.-C. Steinhausen (1), L. Jäncke (2), R. Drech-
sler (1) & D. Brandeis (1,5,6), (1) Departmentof Child and Adolescent Psychiatry, Univer-sity of Zürich, Zürich, Switzerland, (2) Depart-ment of Neuropsychology, Institute for Psy-chology, University of Zürich, Zürich, Switzer-land, (3) Department of Child and AdolescentPsychiatry, University of Erlangen-Nürnberg,Germany, (4) Heckscher-Klinikum, München,Germany, (5) Department of Child and Ado-lescent Psychiatry and Psychotherapy, Cen-tral Institute of Mental Health, Mannheim, Ger-many, (6) Center for Integrative Human Phys-iology, University of Zürich, Zürich, Switzer-land.
In an ongoing study tomographic neuro-feedback (NFB) is evaluated to improvetreatment of attention deficit hyperactivitydisorder (ADHD). Children with ADHD aretrained over 18 sessions (36 units) in twelveweeks to regulate their brain activity (EEGfrequencies and slow cortical potentials) inthe anterior cingulate cortex (ACC). Thirty-one-channel electroencephalogram (EEG)is used to calculate low-resolution electro-magnetic tomographic (sLORETA) NFB. Alltraining sessions start with a baseline restingEEG, which is analyzed for spectral powertomography focusing on theta and beta fre-quency bands. We hypothesized that regionspecific NFB training leads to a continuousdecrease of theta/beta ratios, reflecting an in-creasingly attentive resting EEG in the targetregion across training sessions. Preliminaryanalysis of 5 children (age: 10-13 years) showa decrease of theta/beta ratio in the ACC overthe time course of the whole training. Linearregression is significant across the trainingsessions for the group mean, and individualregression slopes tend to be negative across
54 Human Cognitive Neurophysiology 2010, 3 (1)

W. Skrandies — Abstracts of the 18th German EEG/EP Mapping Meeting
subjects. These results suggest that tomo-graphic NFB helps normalising estimatedintracerebral activity in a specific brain regionaffected in ADHD.Supported by the SBF COST B27 ENOC and by a
grant to the GD of the Kanton of Zürich
2010, 3 (1) Kognitive Neurophysiologie des Menschen 55

Announcements — Ankündigungen
Announcements — Ankündigungen
• ISBET Meeting
The annual meeting of the International Society for Brain Electromagnetic Topography (IS-BET) will take place as a Joint Meeting of ECNS / ISBET / ISNIP in Istanbul, Turkey fromSeptember 14 to 18, 2010.
Information and Registration at: http://www.ecns2010.com/
• Deutsches EEG/EP Mapping Meeting (DMM)
Das 19. Deutsche EEG/EP Mapping Meeting findet vom 15. bis 17. Oktober 2010 inSchloss Rauischholzhausen statt.
Angenommen werden freie Beiträge zu Themen der hirnelektrischen Aktivität des Men-schen in Form von Vorträgen oder Postern.
Schwerpunkte
– Professor Dr. Silvana Galderisi (Napoli) ”The renascence of the electrophysiologicalmethods in schizophrenia research”
– Workshop Dr. Thomas Koenig (Bern) ”Tutorial zur Datenanalyse mit Ragu (RAndom-ization Graphical Userinterface)”
Information und Anmeldung unter: http://www.med.uni-giessen.de/physio/
56 Human Cognitive Neurophysiology 2010, 3 (1)

















