HELMHOLTZ-ZENTRUM POTSDAM2DEPARTMENT OF ARCTIC AND MARINE BIOLOGY, UNIVERSITY OF TROMSØ,
NORWEGEN
Microorganisms are the drivers of the carbon dynamics in permafrostecosystems and the associated release of greenhouse gases. The studyof microbial processes and the underlying microbial communities in Arctic permafrost in consideration of climatic and geochemical data isthe basis for understanding the role of permafrost landscapes in the global climate system and its future development as a carbon source orsink in the scope of a warming Earth.
ó Die arktischen Permafrost-Gebiete spielenaus zweierlei Gründen eine Schlüsselrolle imKlimasystem unserer Erde: Zum einen zeigenBeobachtungen, dass die globale Erwärmungin den hohen Breiten am stärksten ausgeprägtist. Zum anderen ist ein Drittel des globalenKohlenstoffs in Ökosystemen der nördlichenBreiten gespeichert. Um ein besseres Ver-ständnis der gegenwärtigen und zukünftigen
Entwicklung der Kohlenstoffdynamik in kli-maempfindlichen Permafrost-Ökosystemen zuerlangen, konzentriert sich die derzeitige For-schung zur Permafrost-Mikrobiologie aufUntersuchungen zum Kohlenstoffumsatzdurch Mikroorganismen, die Aktivität undStruktur der beteiligten Mikroorganismen-Gemeinschaften und ihre Reaktion auf sichändernde Umweltbedingungen [1].
Die ewige GefrornisDauerfrost beeinflusst mehr als ein Viertelder Erdoberfläche, insbesondere in der nörd-lichen Hemisphäre, sowie Teile der flachenSchelfmeere. Permafrost sind Böden, Sedi-mente oder Gesteine, die mindestens zweiJahre in Folge Temperaturen unter demGefrierpunkt aufweisen. Dabei kann der Dauerfrost mehr als 1.000 Meter in den Unter-grund reichen. Die arktischen Permafrost-Regionen sind durch sehr niedrige Tempera-turen (durchschnittlich –15 °C; Tmin: –48 °C;Tmax: 26 °C) und geringe Niederschlagsmen-gen (90 bis 350 Millimeter pro Jahr) gekenn-zeichnet. Während des kurzen arktischenSommers taut lediglich der oberflächennaheBereich (wenige Zentimeter bis zwei Meter)der Böden auf, der als saisonale Auftauzone(active layer) bezeichnet wird (Abb. 1A). Per-mafrost ist durch Frostmusterbildung (z. B.Polygone; Abb. 1A und C) und verschiedeneandere kryo gene Strukturen gekennzeichnet:Eiskeile (Abb. 1D) sind massive Vereisungs-strukturen, die durch ihr Wachstum über Jahr-hunderte zur Ausbildung der polygonalenFrostmuster in arktischen Tundren führen.Als Taliks (Abb. 1A) werden ungefrorene Sedi-mentschichten im Permafrost bezeichnet, diemeistens unter Wasserkörpern zu finden sind.Cryopegs (Abb. 1A) sind kryogene Struktu-ren im Dauerfrost, die bedingt durch hoheSalzgehalte (30 bis 140 Gramm pro Liter)ungefrorene „flüssige“ Bereiche darstellen.In Cryopegs konnte eine kälte- und salzlie-bende mikrobielle Gemeinschaft nachgewie-sen werden, die an extrem niedrige Nähr-stoffkonzentrationen angepasst ist [2]. Durchdie verschiedenen kryogenen Strukturen wer-den die Lebensbedingungen kleinräumig imÖkosystem bestimmt.
Methankreislauf in Permafrost-BödenDie feuchten Niederungsgebiete der arkti-schen Permafrost-Landschaften sind natürli-che Quellen des klimarelevanten SpurengasesMethan, das neben Wasserdampf, Kohlen-dioxid und anderen Spurengasen unser Erd-klima „anheizt“. Durch die Degradation desPermafrosts im Zuge der Klimaerwärmungwerden große Mengen an organischem Koh-lenstoff, der zuvor in den gefrorenen Sedi-menten weitgehend konserviert war, mikro-
Permafrost-Mikrobiologie
Heiße Prozesse in kalten Böden
˚ Abb. 1: A, Typische Formen der periglazialen Landschaft. B, Permafrost-Boden. C, low-centre-Polygone. D, Eiskeil unter dem Polygonwall. E, Eiskomplex mit Torfeinschlüssen.
biell zugänglich gemacht (Abb. 1E). Der Koh-lenstoffvorrat im Permafrost wird auf bis zu1.650 Gigatonnen geschätzt [3].
Unter anaeroben Bedingungen wird dieserKohlenstoff im Zuge der Methanogenese alsterminaler Schritt am Ende einer Sequenzvon hydrolytischen und fermentativen Pro-zessen durch methanogene Archaeen (Domä-ne Euryarchaeota) zu Methan umgesetzt. Phy-logenetische Analysen zeigen eine hohe Diver-sität dieser Mikroorganismengruppe in dersaisonalen Auftauzone und im darunter lie-genden Permafrost. Die Umweltsequenzenbilden dabei spezifische Permafrost-Cluster,die mit den Familien Methanobacteriaceae,Methanomicrobiaceae, Methanosarcinaceaeund Methanosaetaceae verwandt sind [4]. Diemethanogenen Archaeen in diesen Clusternscheinen ein spezifisches physiologischesPotenzial zu präsentieren, das ihr Überlebenunter den gegebenen extremen Habitat -bedingungen begünstigt. Diese Anpassungwird auch durch die Methanemissionsraten
widergespiegelt, die trotz des extremen sai-sonalen Wechsels von Frieren und Tauenzumindest im arktischen Sommer mit denenaus Feuchtgebieten gemäßigter Breiten ver-gleichbar sind [5].
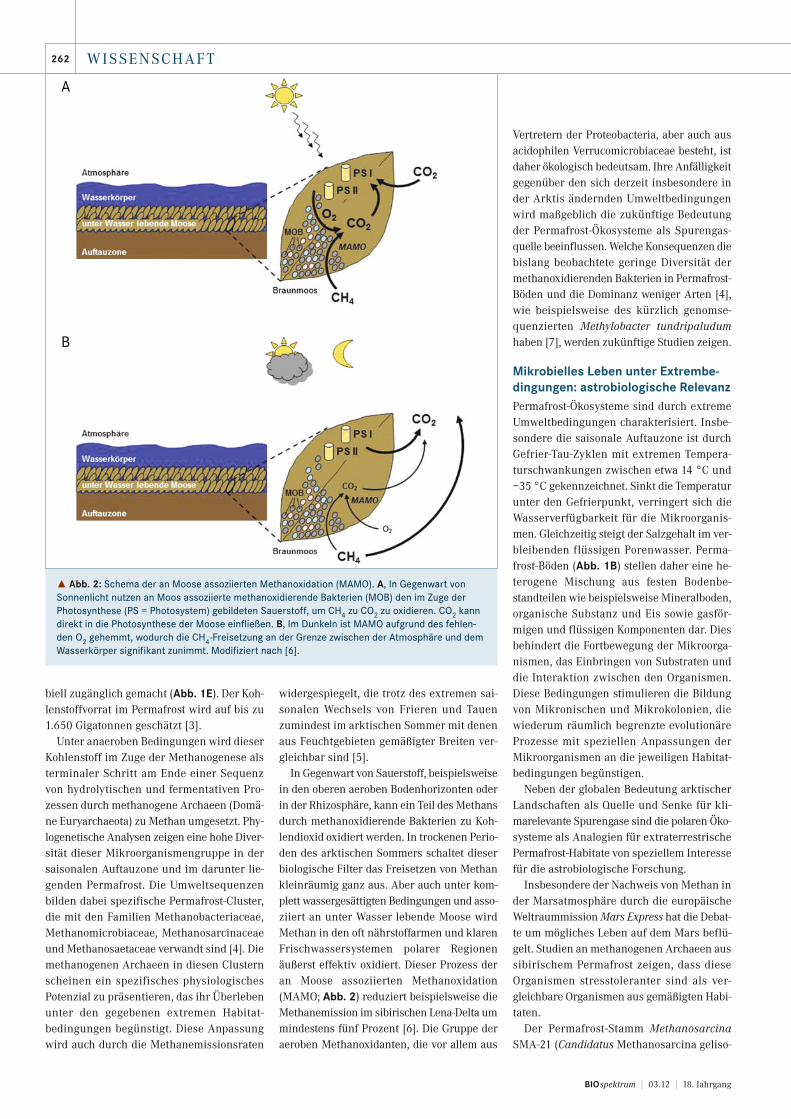
In Gegenwart von Sauerstoff, beispielsweisein den oberen aeroben Bodenhorizonten oderin der Rhizosphäre, kann ein Teil des Methansdurch methanoxidierende Bakterien zu Koh-lendioxid oxidiert werden. In trockenen Perio-den des arktischen Sommers schaltet dieserbiologische Filter das Freisetzen von Methankleinräumig ganz aus. Aber auch unter kom-plett wassergesättigten Bedingungen und asso-ziiert an unter Wasser lebende Moose wirdMethan in den oft nährstoffarmen und klarenFrischwassersystemen polarer Regionenäußerst effektiv oxidiert. Dieser Prozess deran Moose assoziierten Methanoxidation(MAMO; Abb. 2) reduziert beispielsweise dieMethanemission im sibirischen Lena-Delta ummindestens fünf Prozent [6]. Die Gruppe deraeroben Methanoxidanten, die vor allem aus
Vertretern der Proteobacteria, aber auch ausacidophilen Verrucomicrobiaceae besteht, istdaher ökologisch bedeutsam. Ihre Anfälligkeitgegenüber den sich derzeit insbesondere inder Arktis ändernden Umweltbedingungenwird maßgeblich die zukünftige Bedeutungder Permafrost-Ökosysteme als Spurengas-quelle beeinflussen. Welche Konsequenzen diebislang beobachtete geringe Diversität dermethanoxidierenden Bakterien in Permafrost-Böden und die Dominanz weniger Arten [4],wie beispielsweise des kürzlich genomse-quenzierten Methylobacter tundripaludumhaben [7], werden zukünftige Studien zeigen.
Mikrobielles Leben unter Extrem be -dingungen: astrobiologische RelevanzPermafrost-Ökosysteme sind durch extremeUmweltbedingungen charakterisiert. Insbe-sondere die saisonale Auftauzone ist durchGefrier-Tau-Zyklen mit extremen Tempera-turschwankungen zwischen etwa 14 °C und–35 °C gekennzeichnet. Sinkt die Temperaturunter den Gefrierpunkt, verringert sich dieWasserverfügbarkeit für die Mikroorganis-men. Gleichzeitig steigt der Salzgehalt im ver-bleibenden flüssigen Porenwasser. Perma-frost-Böden (Abb. 1B) stellen daher eine he -te ro gene Mischung aus festen Bodenbe-standteilen wie beispielsweise Mineralboden,organische Substanz und Eis sowie gasför-migen und flüssigen Komponenten dar. Diesbehindert die Fortbewegung der Mikroorga-nismen, das Einbringen von Substraten unddie Interaktion zwischen den Organismen.Diese Bedingungen stimulieren die Bildungvon Mikronischen und Mikrokolonien, diewiederum räumlich begrenzte evolutionäreProzesse mit speziellen Anpassungen derMikroorganismen an die jeweiligen Habitat-bedingungen begünstigen.
Neben der globalen Bedeutung arktischerLandschaften als Quelle und Senke für kli-marelevante Spurengase sind die polaren Öko-systeme als Analogien für extraterrestrischePermafrost-Habitate von speziellem Interessefür die astrobiologische Forschung.
Insbesondere der Nachweis von Methan inder Marsatmosphäre durch die europäischeWeltraummission Mars Express hat die Debat-te um mögliches Leben auf dem Mars beflü-gelt. Studien an methanogenen Archaeen aussibirischem Permafrost zeigen, dass dieseOrganismen stresstoleranter sind als ver-gleichbare Organismen aus gemäßigten Habi-taten.
Der Permafrost-Stamm MethanosarcinaSMA-21 (Candidatus Methanosarcina geliso-
˚ Abb. 2: Schema der an Moose assoziierten Methanoxidation (MAMO). A, In Gegenwart vonSonnenlicht nutzen an Moos assoziierte methanoxidierende Bakterien (MOB) den im Zuge derPhotosynthese (PS = Photosystem) gebildeten Sauerstoff, um CH4 zu CO2 zu oxidieren. CO2 kanndirekt in die Photosynthese der Moose einfließen. B, Im Dunkeln ist MAMO aufgrund des fehlen-den O2 gehemmt, wodurch die CH4-Freisetzung an der Grenze zwischen der Atmosphäre und demWasserkörper signifikant zunimmt. Modifiziert nach [6].
A
B
262 WISSENSCHAFT
BIOspektrum | 03.12 | 18. Jahrgang
BIOspektrum | 03.12 | 18. Jahrgang
lum), nahe verwandt mit Methanosarcinamazei, ist beispielsweise durch eine außer-gewöhnliche Toleranz gegen extrem niedri-ge Temperaturen (–78 °C), hohe Salzgehal-te (bis sechs Mol NaCl), Hungern, Aus-trocknung sowie UV- und ionisierendeStrahlung charakterisiert [8]. Dieser Orga-nismus übersteht sogar die thermo-physi-kalischen Bedingungen auf dem Mars, wieein mehrwöchiges Simulationsexperimentzeigte [9]. Daher werden methanogeneArchaeen aus terrestrischem Permafrost alsModelle für die Untersuchung möglichenLebens in extraterrestrischem Permafrost,wie beispielsweise auf dem Mars, genutzt.
Künftige Aufgaben für diePermafrost-MikrobiologieDer Begriff Permafrost-Mikrobiologie unddie damit einhergehende Abgrenzung eineseigenständigen Forschungsfeldes sind nochsehr jung und resultieren vor allem aus derBedeutung von Permafrost-Ökosystemenund ihrer mikrobiellen Gemeinschaft fürdie globale Spurengasbilanz. Inwieweit diemikrobielle Permafrost-Gemeinschaft inihrer Struktur, vor allem aber in ihrem funk-tionellen und physiologischen Repertoireeinzigartig ist und ob sie sich in ihrer Ver-letzlichkeit grundsätzlich von anderenmikrobiellen Gemeinschaften aus bei-spielsweise weniger extremen Habitatenunterscheidet, ist bislang unklar. Kompeti-tive und genombasierte Studien von Per-mafrost-Isolaten können hierzu zukünftigebenso Aufschluss geben wie Simulations-experimente unter definierten Umweltbe-dingungen in ungestörten Bodenkörpern(Mesokosmen) in Kombination mit Metage-nomik und Metatranskriptomik. ó
Literatur[1] Graham DE, Wallenstein MD, Vishnivetskaya TA et al.(2011) Microbes in thawing permafrost: the unknown varia-ble in the climate change equation. The ISME J 6:709–712[2] Gilichinsky DA, Rivkina E, Bakermans C et al. (2005)Biodiversity of cryopegs in permafrost. FEMS Microbiol Ecol53:117–128[3] Tarnocai C, Canadell JG, Schuur EAG et al. (2009) Soilorganic carbon pools in the northern circumpolar perma-frost region. Global Biogeochem Cyc 23:GB2023[4] Vishnivetskaya TA, Liebner S, Wilhelm R et al. (2012)Microbial carbon cycling. In: Miller RV, Whyte LG (Hrsg)Polar Microbiology: Life in a Deep Freeze. ASM Press,Washington, DC. 183–199[5] Parmentier FJW, van Huissteden J, van der Molden KMet al. (2011) Spatial and temporal dynamics in eddy covari-ance observations of methane fluxes at a tundra site in nor-theastern Siberia. J Geophys Res 116:G03016[6] Liebner S, Zeyer J, Wagner D et al. (2011) Methane oxi-dation associated with submerged brown mosses reducesmethane emissions from Siberian polygonal tundra. J Ecol99:914–922[7] Svenning MM, Hestnes AG, Wartiainen I et al. (2011)Genome sequence of the Arctic methanotrophMethylobacter tundripaludum SV96. J Bacteriol 193:6418–6419[8] Morozova D, Wagner D (2007) Stress response ofmethanogenic archaea from Siberian permafrost comparedto methanogens from non-permafrost habitats. FEMSMicrobiol Ecol 61:16–25[9] Morozova D, Möhlmann D, Wagner D (2007) Survival ofmethanogenic archaea from Siberian permafrost undersimulated Martian thermal conditions. Origin of Life andEvolution of Biospheres 37:189–200
Dr. Susanne LiebnerUniversity of TromsøDepartment of Marine and Arctic BiologyFakturamottakNO-9037 TromsøTel.: +47-(0)776-23318Fax: +47-(0)[email protected]
AUTORENDirk WagnerJahrgang 1963. 1998 Promotion in Bodenmikrobiologie an der Universität Ham-burg. 2002–2012 Aufbau und Leitung der AG Geomikrobiologie in Periglazialgebie-ten am Alfred-Wegener-Institut, Potsdam. 2007 Habilitation am Institut für Bioche-mie und Biologie der Universität Potsdam. Seit 2012 Leiter der Sektion Geomikro-biologie am Helmholtz-Zentrum Potsdam, Deutsches GeoForschungsZentrum undProfessor für Geomikrobiologie der Universität Potsdam.
Susanne LiebnerJahrgang 1978. 2003 Diplom in Geoökologie, Universität Potsdam. 2005 Master ofScience (Marine Mikrobiologie), International Max Planck Research School (Mar-Mic). 2008 Promotion in der Gruppe Geomikrobiologie am Alfred-Wegener-Institut,Potsdam. Seit 2008 Postdoc, zunächst in der Gruppe Umweltmikrobiologie, ETH Zürich, Schweiz, und anschließend seit 2010 an der Universität Tromsø, Norwegen,am Institut für Arktische und Marine Biologie.