Institut fur Angewandte Botanik der Universitat Hamburg Der EinfluB phenolischer Aktivatoren und Inhibitoren der IES-Oxydase-Aktivitat auf die Adventivbewurzelung bei Tradescantia albiflora The Influence of Phenolic Promotors and Inhibitors of IAA-Oxidase-Activity on the Induction of Adventitious Roots in Tradescantia albiflora UWE PINGEL Mit 1 Abbildung Eingegangen am 18. Dezember 1975 . Angenommen am 20. Februar 1976 Summary Phenolic promotors and inhibitors of IAA-oxidase actiVIty stimulate the induction of adventitious roots in Tradescantia. The former are physiologically more effective with the exception of homovanillic acid, which is the strongest inductor although it inhibits IAA- oxidase activity. The growth of the adventitious roots is, on the other hand generally inhib- ited by all phenolic compounds. Some of these applied phenolic acids are native substances of the cuttings. Correlative to root promotion and the effect on IAA-oxidase activity is the increase of oxygen uptake by the nodes of Tradescantia after phenolic treatment. The relationship of this effect and of root induction to the uncoupling of oxidative phosphory- la tion is discussed. Key words: phenolic substances, IAA-oxidase activity, adventitious roots, Tradescantia. Einleitung Die Regeneration von Wurzeln an Stecklingen wird heute vielfach durch Applikation von «Bewurzelungshormonen» wie IES, IBS und NES unterstiitzt. Trotz dieser erfolgreichen Anwendung in der Praxis sind die Vorstellungen iiber den Ablauf des Induktionsmechanismus noch weitgehend unklar. Ausgehend von der Diskussion des Rhizokalinkomplexes (BOUILLENNE u. WENT, 1933) sieht LIBBERT (1956) die ersten Teilschritte einer Induktion von Adventivwurzeln in einer Komplexbildung der IES mit einem mobilen Faktor X, der durch o-diphenolische Gruppen charakterisiert wird. Durch Vergleich von Extrakten leicht und schwer zu bewurzelnder Pflanzen kommt HESS (1965 a, 1968) zu dem Schlug, dag synergistisch mit der IES wirkende Co-Faktoren (an Orten spezifischer Phenolaseaktivitat) die Bewurzelungsfahigkeit steuern. z. P/lanzenphysiol. Ed. 79. S. 109-120. 1976.
Transcript
Institut fur Angewandte Botanik der Universitat Hamburg
Der EinfluB phenolischer Aktivatoren und Inhibitoren der IES-Oxydase-Aktivitat auf die Adventivbewurzelung bei Tradescantia albiflora
The Influence of Phenolic Promotors and Inhibitors of IAA-Oxidase-Activity on the Induction of Adventitious Roots in Tradescantia albiflora
UWE PINGEL
Mit 1 Abbildung
Eingegangen am 18. Dezember 1975 . Angenommen am 20. Februar 1976
Summary
Phenolic promotors and inhibitors of IAA-oxidase actiVIty stimulate the induction of adventitious roots in Tradescantia. The former are physiologically more effective with the exception of homovanillic acid, which is the strongest inductor although it inhibits IAAoxidase activity. The growth of the adventitious roots is, on the other hand generally inhibited by all phenolic compounds. Some of these applied phenolic acids are native substances of the cuttings. Correlative to root promotion and the effect on IAA-oxidase activity is the increase of oxygen uptake by the nodes of Tradescantia after phenolic treatment. The relationship of this effect and of root induction to the uncoupling of oxidative phosphoryla tion is discussed.
Die Regeneration von Wurzeln an Stecklingen wird heute vielfach durch Applikation von «Bewurzelungshormonen» wie IES, IBS und NES unterstiitzt. Trotz dieser erfolgreichen Anwendung in der Praxis sind die Vorstellungen iiber den Ablauf des Induktionsmechanismus noch weitgehend unklar.
Ausgehend von der Diskussion des Rhizokalinkomplexes (BOUILLENNE u. WENT, 1933) sieht LIBBERT (1956) die ersten Teilschritte einer Induktion von Adventivwurzeln in einer Komplexbildung der IES mit einem mobilen Faktor X, der durch o-diphenolische Gruppen charakterisiert wird. Durch Vergleich von Extrakten leicht und schwer zu bewurzelnder Pflanzen kommt HESS (1965 a, 1968) zu dem Schlug, dag synergistisch mit der IES wirkende Co-Faktoren (an Orten spezifischer Phenolaseaktivitat) die Bewurzelungsfahigkeit steuern.
z. P/lanzenphysiol. Ed. 79. S. 109-120. 1976.
110 U. PINGEL
In diesen Modellen besitzen best. phenolische Subs tan zen eine regelnde Funktion bei der Wurzelinduktion. Speziell auf dem Antagonismus zwischen Phenolcarbonsauren (peS) yom o-dihydroxy-Typ und p-hydroxylierten Phenolen im Zusammenhang mit der Auxinapplikation liegt in der Diskussion das Schwergewicht. Die Ursache dieses gegensatzlichen Einflusses auf die Induktion sieht man in der antagonistischen Wirkung der Phenole auf die Aktivitat des IES-Oxydase-Systems (TOMASZEWSKI u. THIMANN, 1966; SHANTZ, 1966).
Nach Befunden von TULI u. MOYED (1969), BASU u. TULI (1972) solI ein Oxydationsprodukt der IES - das im Vergleich mit dem genuinen Auxin physiologisch starkere Methylenoxindol - als der Effektor des IES-vermittelten Wachs turns agieren. Spielt nun das Methylenoxindol auch bei der Adventivwurzelinduktion die Rolle des Effektors, wie es die Basisakkumulation von Metaboliten der IES in Stecklingsversuchen von GREENWOOD und GOLDSMITH (1970) andeuten konnte, so ware eine Abhangigkeit der Bewurzelungsrate yom Spiegel der IES-Oxydationsprodukte gegcben. Der IES-Oxydase kame damit (entgegen ihrer urspriinglichen Rolle) eine positive Funktion zu.
Zur Regelung der Enzymaktivitat wurde deshalb mit phenolischen Aktivatoren und Inhibitoren gearbeitet, urn deren EinfluB auf die Adventivwurzelinduktion -als Ausdruck der Steuerung des Methylenoxindolniveaus - zu charakterisieren.
Da aber diese Phenole auch in weitere fundamentale Prozesse, wie die der ATP-Regeneration und Respiration, eingreifen (LIEBERMANN u. BIALE 1956, DEDONDER u. VAN SUMERE 1971, KEFELI u. KADYROV 1971), diese zugleich Induktionsvorgange beriihren, wie die Steigerung der Adventivbewurzelung durch 2,4DNP (RuGE 1969) gezeigt hat, sol1te ebenfa11s auf dieser allgemeinen Ebene die Wirkung der Phenolapplikation kontrolliert werden (Messung der A tmungsintensi tat).
Material und Methoden
Ais Versuchsobjekt dienten Stecklinge eines Klons von Tradescantia albiJlora Kunth. Die Tradescantienstecklinge zeichnen sich durch eine leichte Regeneration des Wurzelsystems aus. Gewinnung und Haltung der Stecklinge siehe RUGE (1969) und BRUNNER (1970).
Fiir die Versuche wurden folgende PCS in waBriger Lasung benutzt: p-Cumarsaure (Cum), Kaffeesaure (Kaf), Ferulasaure (Fer); p-OH-Benzoesaure (P-B2S),
Die Auswertung der stets 10tagigen Bewurzelungsversuche (Einwirkungsdauer der Versuchslasungen 1, 3 und 10 d) bezog sich auf die Adventivwurzelanlagen (WA), die freien Wurzeln (W) sowie auf die durchschnittliche EinzelwurzelHinge (Wd.
Die Variation in der Gesamtbewurzelungsrate (WA + W) gibt der Differenzwert W A + W (Versuch) - W A + W (KontroUe) an. Die Daten aUer Versuchsglieder wurden dabei in % der KontroUe (= 100) ausgedriickt. Je Versuchsgruppe betrug die Anzahl der Stecklinge 40 bzw. 20. Bei 2maliger Wiederholung (n-gesamt = 100) bedeutete dies eine gute statistische Absicherung: + bedeutet Signifikanz auf dem 5-, + + auf dem 1-, + + + auf dem 0,1-Prozent-Niveau; - bedeutet keine Signifikanz vorhanden.
z. PJlanzenphysiol. Ed. 79. S. 109-120. 1976.
Phenole und Adventivwurzel-Bildung bei Tradescantia 111
Zur Identifizierung nativer Phenolcarbonsauren von Tradescantia und eventuell met abolisierter applizierter Phenole wurden dlinnschichtchromatographische Verfahren angewandt (STAHL, 1967). Das Ausgangsmaterial flir diese Untersuchungen bildeten die 5.-7. Knoten mit N2 tiefgefrorener Stecklinge. Der nach RIBEREAU-GAYON (1973) veranderte und gewonnene Extrakt wurde (zur Bestimmung der gebundenen PCS) auch einer alkalischen und aciden Hydrolyse unterworfen.
Auf saurestabilen Kieselgel G Fertigfolien und Celluloseplatten wurde mit verschiedenen Laufmitteln (STAHL, 1967; VAN SUMERE et a!., 1965) getrennt. Die Identifizierung der PCS erfolgte mit Hilfe reiner Vergleichssubstanzen, der Fluorescenz im UV-Licht sowie durch Anfarbung mit dem Diazotierungsreagenz Benzidin.
Die Bestimmung der Atmungsintensitat wurde mit einem Warburg-Gerat bei 20 DC und liber 2 h durchgeflihrt. Getestet wurden die 6. Knoten von Stecklingen, die flir 24 bzw. 72 h mit der Versuchslosung behandelt worden waren. Je 10 solcher Nodi dienten zur Beschickung eines AtmungsgefaBes. Da die Nodi nod! Chlorophyll enthielten, fan den die Versuche im Dunkeln statt.
Die 02-Aufnahme und die CO2-Abgabe der Knoten wurde in ftllg Frischgewicht ausgedrlickt und auf die Kontrolle bezogen.
Untersuchungen und Ergebnisse
Bewurzelungsversuche
In der genannten Versuchsanstellung konnen allgemein 3 Entwicklungsphasen unterschieden werden: Die Induktion und Ausdifferenzierung einer Wurzelanlage (W A), das Auswachsen zu einem funktionsfahigen Organ (W) und das Uingenwachstum der Wurzeln (WL). Die Induktionsfahigkeit wird durch die beiden ersten Prozesse (W A + W) ausgedriickt. Tabelle 1 gibt eme Obersicht der Bewurzelungsergebnisse.
Zimtsaurederivate: Durch Kaffee- und Ferulasaure tritt nach einer Behandlung bis zu 10-4 mol einschlieBlich keine signifikante Anderung in der Bewurzelungsrate ein. Mit zunehmender Konzentration und fortschreitender Behandlungsdauer erhoht sich dann die Gesamtzahl der Adventivwurzelanlagen. Ein Maximum wird unter diesen Versuchsbedingungen - wie auch bei den iibrigen Phenolen - nicht iiberschritten ; die Steigerungsrate laBc aber zu hohen Konzentrationen und langeren Einwirkungszeiten hin nacho
Der Anteil der freien Wurzeln im Verhaltnis zur Gesamtwurzelanzahl {w2:w} sinkt mit zunehmender Versuchdauer und steigender Molaritat der Losung ebenfalls abo Das ist eine auch bei den anderen Phenol en zu beobachtende gesetzmaBige Erscheinung, da die durch die Testsubstanz zum Versuchsende induzierten Anlagen
nicht mehr auszuwachsen vermogen. P-Cumarsaure zeigt dagegen eine wesentlich intensivere Stimulierung der
Bewurzelung, die bei 2 . 10-3 mol iiber 10 Tage ca. 165 % der Kontrolle betragt. Da aber Kaf und Fer als bewahrte Hemmer der IES-Oxydase gelten (VARGA U.
KavEs, 1962; SACHER, 1963), wahrend Cum als starker Forderer beschrieben wird (KONINGS, 1964; LEE u. SKOOG, 1965; PILET et aI., 1967), stimulieren sowohl
Z. P/lanzenphysiol. Bd. 79. S. 109-120. 1976.
112 U. PINGEL
Aktivatoren als auch Inhibitoren die Adventivwurzelinduktion. Erstere erweisen sich dabei als physiologisch wirksamer.
Benzoesaurederivate: Prot und Van weisen deutliche Parallel en zu den entsprechenden Zimtsaurederivaten auf. Die Steigerung der Bewurzelungsrate fallt aber bei Prot gegenuber Kaf zuruck. Die Stimulierung durch p-Bzs erfahrt nur bei hochster Konzentration und Applikationszeit eine uberdurchschnittliche Steigerung, ohne da6 die Ausbildung Freier Wurzeln in gleicher Weise gefordert wird. Auch gibt es fur die p-Bzs neben Befunden einer IES-Oxydase-Aktivitatsforderung (PI LET, 1966; JANSSEN, 1969) auch solche einer neutralen Reaktion (BASU 1970). Die Differenz im Bewurzelungsverhalten durch Aktivatoren und Inhibitoren (Prot und Van: ABERG u. JOHANSSON, 1969; JANSSEN, 1970) ist hier also nicht so ausgepragt.
Phenylessigsaurederivate: Der absolut geringe Anstieg der Bewurzelung durch 3,4 Pes, der sich nur bei starkster Konzentration und 10tagiger Applikation signifikant von der Kontrolle unterscheidet, kann mit der extremen Anfalligkeit gegen Oxydantien interpretiert werden. Dies fuhrt zur Braunung und Gewebeschadigung in der Stecklingsbasis. Daher sinkt hier ebenfalls der Anteil funktionsfahiger Wurzeln pro Steckling.
Dagegen ist die Hom hinsichtlich der Induktion mit den anderen Methoxyderivaten nicht mehr vergleichbar, da schon nach 3tagiger Applikation und 10-4 mol eine signifikante Erhohung der Bewurzelungsrate erreicht wird, die sich bei 2.10-3 mol und 10 d auf nahezu 100 % steig ern la6t. Sie ist damit signifikant gro6er als die Induktion durch das p-hydroxy-Derivat.
Die Induktionsverhaltnisse der beiden vorigen Gruppen lassen sich also nicht auf die Pes-Derivate ubertragen, da der Inhibitor Hom (GORTNER u. KENT, 1958) den Aktivator p-Pes (RUNKOVA et aI., 1972) in der Bewurzelungsrate ubertrifft.
Durchschnittliche Einzelwurzellange
Das Langenwachstum der freien Wurzeln wird generell bei 10tagiger Behandlung und 10-4 mol - hier wird die Induktion noch nicht tangiert - signifikant gehemmt. Steigende Konzentrationen verringern die Wurzellange je nach der eingesetzten pes stetig bis auf maximal 5-24 Ufo. Allgemein gilt dies jedoch nur bei Applikation uber den vollen Versuchszeitraum, da die Wurzeln dann selbst an der Aufnahme der Phenole beteiligt sind, wahrend bei 1- und 3tagiger Behandlung von den in den Nodi akkumulierten (und nur uber die Schnittflache aufgenommen) Phenol en noch keine inhibierende Wirkung auf das spater einsetzende Wachs tum der freien Wurzeln ausgeht.
Dabei zeigt sich, da6 die IES-Oxydase-Inhibitoren mit einer maximalen Reduktion (bis 14-24 Ufo der Kontrolle) signifikant schwacher wirken als die zugehorigen Aktivatoren (6-13 Ufo). Eine Ausnahme bildet hier wieder die Hom, welche von dies em Schema abweichend bei nur 3tagiger Applikation und 2.10-3 mol schon eine starke inhibitorische Wirkung ausubt. Diese Hemmung la6t dar auf schlie6en, da6
z. Pjlanzenphysiol. Bd. 79. S. 109-120. 1976.
Phenole und Adventivwurzel-Bildung bei Tradescantia 113
bereits schwache Konzentrationen, wahrend der Differenzierungsphase in den Wurzel anlagen gespeichert, das spatere Langenwachstum der Wurzeln verringern.
Chromatographische Untersuchungen
Da durch einen einfachen Oxydationsschritt - z. B. vermittelt durch das Phenolasesystem - im Steckling die Mono- zu Diphenolen umgesetzt werden konnen und damit auch ihre Funktion als Regulatoren umkehren, war es notwendig, die hier applizierten Phenole auf diesen Metabolismus hin zu untersuchen. Das war nur moglich im Anschlug an eine qualitative Analyse nativer Phenolcarbonsauren, urn Verwechselungen applizierter mit endogener PCS zu vermeiden.
1m unhydrolysierten bzw. acid hydrolysierten Extrakt der Nodi frisch geschnittener oder als Kontrollstec.ldinge gehaltener Tradescantien wurden keine freien Phenolcarbonsauren gefunden. Dagegen konnten im Chromatogramm eines alkalisch hydrolysierten Extraktes in abnehmender Konzentration Ferula-, Kaffee-, Cumar- und Sinapinsaure nachgewiesen werden.
Diese bei Tradescantia endogen in den Knoten als Orten der Wurzelinduktion vorliegenden Phenole gehoren ausschliegJich zu den Zimtsaurederivaten. Nach dem von NEISH (1965) und PRIDHAM (1965) beschriebenen Biosyntheseweg gehen sie stufenweise aus der Cumarsaure hervor, d. h. es entstehen hier aus einem Aktivator als Ausgangsprodukt mehrere Inhibitoren der IES-Oxydase. Diese Phenole sind zwar innerhalb der Pflanze als Ester gebunden, aber nach RABIN u. KLEIN (1957), ENGELSMA (1965) bugen sie ihre Fahigkeit der Enzymregulation nicht ein und konnten so am Induktionsmechanismus beteiligt sein.
Nach 3tagiger Applikation best. Zimtsaurederivate verandert sich das Chromatogramm eines unhydrolysierten Extraktes von Stecklingsknoten: Neben der applizierten Cumarsaure treten jetzt Kaffeesaure und ebenfalls Spuren von Ferulasaure auf (die sich auch nach Zugabe von Kaf nachweisen lagt). Die Herkunft der Fer kann dabei der Spaltung des nativen Esters wahrend der Isolierungsphase entspringen, eventuell aber auch durch weitere Substitution des Intermediarproduktes Kaf entstanden sein. Die strukturelle Anderung der Cumarsaure lauft also auf eine Umkehrung ihrer Wirkung auf die IES-Oxydase hinaus. Geschwindigkeit und Quantitat der Umsetzung und damit die Bedeutung fur die Induktion sind gering.
Die applizierten Benzoe- und Phenylessigsaureabkommlinge bleiben im wesentlichen unmetabolisiert. Ein Vergleich einfacher und hydrolysierter Extrakte gibt keine fagbaren Differenzen. Ausgenommen sind die sehr oxydablen o-Diphenole, die ein Gemisch diffuser Melaninprodukte ergeben. Dies fuhrt nicht zu Funktionsumkehr sondern nur zu Funktionsverlust.
Atmungsuntersuchungen
Nach Applikation verschiedener Phenole treten Veranderungen des Atmungsquotienten nicht auf, wie die Bestimmungen des O2- Verbrauchs und der CO2-
z. PJlanzenphysiol. Bd. 79. S. 109-120. 1976.
114 U. PINGEL
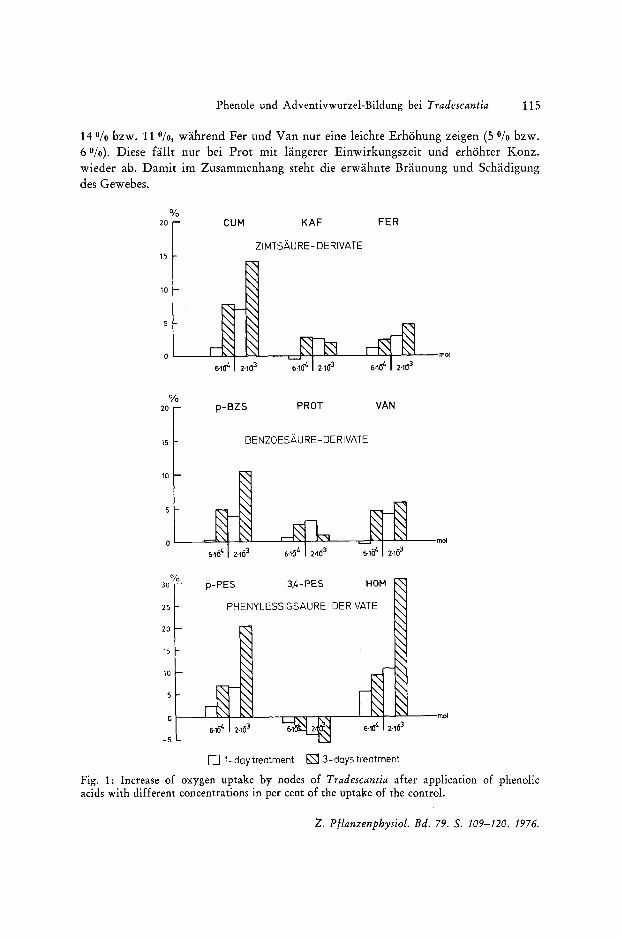
Abgabe ergaben. Die PCS werden also nicht als zusatzliches Respirationssubstrat in den Stoffwechsel miteinbezogen; eine Anderung des O 2 Verbrauchs kann also darauf nicht zuriickgefiihrt werden. Ein Vergleich der 3 phenolischen Gruppen zeigt nun (Abb. 1), dag die Reaktion der Tradescantiaknoten auf Zimt- und Benzoesaurederivate weitgehend parallel lauft. Cum und p-Bzs als starke Forderer der Adventivbewurzelung verursachen einen Anstieg des O 2 Verbrauchs urn maximal
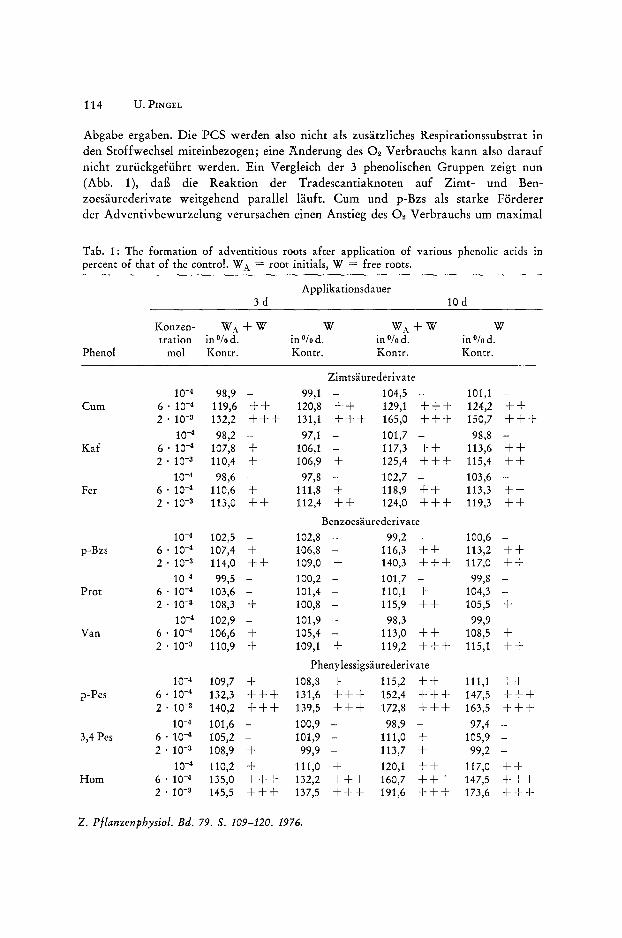
Tab. 1: The formation of adventitious roots after application of various phenolic acids in percent of that of the control. W A = root initials, W = free roots.
Applikationsdauer 3d 10 d
Konzen- WA+W W WA+W W tration in Ofo d. in Ofo d. in Ofo d. in Ofo d.
Phenol mol Kontr. Kontr. Kontr. Kontr.
Zimtsiiurederi va te 10-4 98,9 - 99,1 - 104,5 - 101,1 -
Phenole und Adventivwurzel-Bildung bei Tradescantia 115
14 % bzw. 110/0, wahrend Fer und Van nur eine leichte Erhohung zeigen (5 Ofo bzw. 6 %). Diese fallt nur bei Prot mit langerer Einwirkungszeit und erhohter Konz. wieder abo Damit im Zusammenhang steht die erwahnte Braunung und Schadigung des Gewebes.
% 20
15
10
0/0
20
15
% 30
25
20
15
10
-5
CUM
p-BZS
KAF
ZIMTSAURE-DERIVATE
PROT
BENZOESAURE-DERIVATE
p-PES 3.4-PES
FER
VAN
PHENYLESSIGSAURE- DERIVATE
o 1-daytreatment bS]3-daystreatment
Fig. 1: Increase of oxygen uptake by nodes of Tradescantia after application of phenolic acids with different concentrations in per cent of the uptake of the control.
z. PJlanzenphysiol. Bd. 79. S. 109-120. 1976.
116 U. PINGEL
Wesentlich intensiver ausgepragt ist diese Tendenz der o-Diphenole bei der 3,4 Pes, so daB es dort zu einem einmaligen Absinken der O2 Aufnahme kommt.
Die Ausnahmestellung der Pes-Derivate wird noch durch die Homovanillinsaure (als IES-Oxydase-lnhibitor) betont, die mit ca. 30010 den weitaus groBten Anstieg im O2 Verbrauch verursacht (p-Pes nur ca. 20010). Eine enge Parallele findet das abweichende Verhalten dieser Derivate in den Adventivwurzelergebnissen, die auch bei den anderen Phenolen die Atmungsresultate widerspiegeln.
Diskussion
In nachstehender Tabelle 2 sind die wichtigsten Ergebnisse des experiment ellen Teils dieser Arbeit zusammengestellt. Danach zeigen die in dieser Untersuchung verwendeten Phenolcarbonsauren je nach Konzentration und Applikationsdauer eine schwach bis stark positive Wirkung auf die Neubildung von Adventivwurzeln. Niedere Konz. und kiirzere Einwirkungszeiten bedingen allgemein eine schwachere Forderung dieses Prozesses als hohere Konz. und langere Einwirkungszeiten. Eine Hemmung trat bei keinem der untersuchten Phenole auf.
Innerhalb der einzelnen Stoffgruppen bewirken die p-OH-Derivate eine zwar unterschiedliche, aber generell hohere Bewurzelungsrate als die o-Diphenole bzw. die methoxy-substituierten Abkommlinge. Von diesem Prinzip weicht lediglich die Homovanillinsaure abo Die Sonderstellung dieses nicht biogenen Pes-Derivates im
Tab. 2: Summary of the experimental data.
nat ive Funktions-Adv.wurzel· i nduktion Einflun 5teigerung Phenole wechsel schw. hohe au f 02 bei durch
Konz.
rES Oxydos<?
Ferderer. Hemmer- Wurzellge. Aufnahme Tradesc. Meta bol. Oxydation
Z imtsaurederi v.
p-Cum :!: +++ +
Kaf :!: +
Fe r :!: +
Benzoesciuredec
p- B z s :!: ++ +Etl
Prot :!: +
Van :!: +
Phenylessi gs. der.
p- Pes + +++ +
3.4 - Pes :!: + -? Hom + ++++
Z. PJlanzenphysiol. Bd. 79. S. 109-120. 1976.
+ +
(+) +
(+) +
+
:!: (+)
++
++
+ (+)
(+)
+
++
Phenole und Adventivwurzel-Bildung bei Tradescantia 117
Verhaltnis zu Van und Fer la~t sich vielleicht aus dem Wuchsstoffcharakter der Muttersubstanz ableiten (FRIES, 1968).
Bis auf die Hom entspricht also die unterschiedlich starke Induktion neuer Anlagen in allen Stoffgruppen einer Trennung von IES-Oxydase Aktivatoren und Inhibitoren.
Nach der eingangs formulierten Hypothese sollten die applizierten Aktivatoren ja auch eine Stimulierung der Bewurzelung uber den Methylenoxindoleffektor herbeifuhren. Damit im Einklang stehen Ergebnisse uber die Akkumulation von Komponenten mit multipler Enzymfunktion (PPO, IES-Oxydase, Peroxydase) auf fruhen Stufen der Initialenbildung (FRENKEL u. HESS, 1973). Der genannten Hypothese brauchen aber nicht Befunde zu widersprechen, da~ bei simultaner Versorgung des Stecklings mit exogener IES und p-Phenolen die Adventivbewurzelung nicht gesteigert wird.
Solche Ergebnisse lie~en sich unter der Annahme erklaren, da~ zwar auch in diesen Experimenten der Methylenoxindoleffektor vermehrt gebildet wird, dieser aber bei exogener Zugabe von IES bereits an den Schnittflachen und Orten der Pflanze entsteht, an denen keine Bindung an definiertes Protein (BASU u. TULI, 1972) stattfindet. Somit kann weder eine aktive Phase der IES hergestellt werden noch eine Stabilisation des labilen Oxydationsproduktes erfolgen. Methylenoxindol wird zu inertem Methyloxindol abgebaut (MOYED, 1967) und die IES Wirkung damit gemindert.
Entsprechend konnen auch Versuchsergebnisse interpretiert werden, bei denen durch rein exogene Zufuhr von Methylenoxindol eine Stimulierung der Adventivwurzelinduktion nicht erreicht wurde (ANDERSEN et aI., 1972).
1m Gegensatz zu der genannten Hypothese stehen jedoch die Befunde emer wenn auch wesentlich schwacheren - Steigerung der Induktion durch IES-Oxydase Inhibitoren. Die starkste Stimulation wird allerdings durch die Hom ausgelost, die aus strukturchemischen Grunden und nach GORTNER u. KENT (1958) als IES-Oxydase Inhibitor zu werten ist.
Es mussen also noch andere Eigenschaften der Phenole die Induktion gesteuert oder zumindest die Wirkung auf die IES-Oxydase iiberlagert haben. Z. B. konnte sich der positive Einflu~ der phenolischen Hemmer auf die Bewurzelung uber einen vermehrten Abbau von Tryptophan zu IES durchgesetzt haben, da jene «Inhibitoren» dies en enzymatischen Proze~ beschleunigen (GORDON u. PALEG, 1961; LIBBERT et al. 1969).
Die auffallende Korrelation zwischen Stimulation der Ot Aufnahme und Bewurzelungshohe gibt ebenfalls einen Hinweis auf einen anderen Angriffspunkt der Phenole im Stoffwechsel, der auf einer Forderung des oxydativen Metabolismus basieren konnte (KAMINSKI, 1959; WEISER, 1967).
Es wurde nun aber von RUGE (1969, 1971) gezeigt, da~ eine teilweise Blockierung der Atmungsfermente die Adventivwurzelinduktion nicht wesentlich beeintrachtigt,
z. P/lanzenphysiol. Bd. 79. S. 109-120. 1976.
118 U. PINGEL
aber eine Hemmung der ATP-Synthese durch 2,4 DNP die Anlagenbildung zu steigern vermag. Daher ist die Erhohung der Respirationstatigkeit moglicherweise nur ein sekundarer Effekt einer pnmar entkoppelnden Wirkung auf die oxydative Phosphorylierung (GAUR, 1959; DEDONDER u. VAN SUMERE, 1971; STENLID, 1963).
Wah rend die Neubildung von Adventivwurzelanlagen durch die hier untersuchten Phenole generell gefordert wird, ist das Langenwachstum der Wurzeln im Dauerversuch stets reduziert. Der negative EinfluB auf die Wurzellange lauft im allgemeinen parallel der gesteigerten Anlageninduktion und erhohter O2 Aufnahme. Die Wachstumshemmung setzt aber bereits bei schwachen Konzentrationen ein, die die Induktion noch nicht beriihren. Konzentrationen, die morphologische Prozesse zu modifizieren vermogen, liegen also auch hier hoher als die des Langenwachstums (KEFELI, 1968).
Obwohl bei hohen Phenolkonzentrationen Wundeffekte, die oft einen unspezifischen, positiven EinfluB auf die Bewurzelung haben (SOEKAR]O u. JANSSEN, 1969), eine Rolle spiel en konnen, sind die Phenolwirkungen sicher nicht mit dem bewurzelungssteigernden Effekt sogenannter «injurious substances» an der Schwelle zur Toxicitat zu vergleichen, wie es in den Versuchen von SOEKAR]O (1965) zum Ausdruck kommt.
Der hier untersuchte EinfluB auf die Induktion der Adventivbewurzelung resultiert also mehr aus einer Wechselwirkung mehrerer Eigenschaften der strukturverschiedenen Phenole, die z. T. gegensatzlichen Charakter tragen.
Literatur
ABERG, B., and I. JOHANSSON: Studies on plant growth regulators. XXIV. Some phenolic compounds. Lantbrukshogskolans Annaler 35, 3-27 (1969).
ANDERSEN, A. 5., 1. B. M0LLER, and J. HANSEN: 3-Methylene-oxindole and plant growth regulation. Physiol. Plant. 27, 105-108 (1972).
BASU, R. N.: Indole acetic acid oxidizing system in relation to synergism and antagonism between auxins and non-auxinic chemicals in rooting of cuttings. Indian J. PI. Phys. 13,249-262 (1970).
BASU, P. 5., and V. TULI: Auxin activity of 3-methylene-oxindole in wheat. Plant Phys. 50, 499-502 (1972).
- - The binding of indole-3-acetic acid and 3-methylene oxindole to plant macromolecules. Plant. Phys. 50, 503-506 (1972).
BRUNNER, H.: Beeinflussung der Adventivwurzelbildung bei Tradescantia und Fuchsia durch Marko- und Mikronahrstoffe 1. Makronahrstoffe. Gartenbauwissenschaften 35, 353-364 (1970).
BOUILLENNE, R.: Aspects physiologique de la formation des racines. Bull. Soc. Roy. Belgique 95,193-204 (1964).
BOUILLENNE, R., and R. F. WENT: Recherches experiment ales sur la reformation des racines dans les plantules et les boutures des plantes superieures. Ann. Jard. Bot. Buitenzerg 43, 25-202 (1933).
DEDONDER, A., and C. F. VAN SUMERE: The effect of phenolics and related compounds on the growth and respiration of ChIarella vulgaris. Z. Pflanzenphysiol. 65, 70-80 (1971).
ENGELSMA, G., and G. MEIJER: The influence of light and different spectral regions on the
Z. PJlanzenphysial. Bd. 79. S. 109-120. 1976.
Phenole und Adventivwurzel-Bildung bei Tradescantia 119
synthesis of phenolic compounds in Gherkin seedlings in relation to photomorphogenesis. II. IAA-oxydase activity and growth. Acta Bot. Neerl. 14, 73-92 (1965).
FRENKEL, c., and C. E. HESS: Isozymic changes in relation to root initiation in mung bean. Canad. J. Bot. 52, 295-297 (1973).
FRIES, K.: Phenole und Wachstum I. Beitrage Bio!. d. Pflanzen 44,289-318 (1968). - Phenole und Wachstum II. Beitrage Bio!. d. Pflanzen 45,143-156 (1968). GAUR, B. K., and H. BEEVERS: Respiratory and associated responses of carrot discs to sub
stituted phenols. Plant Phys. 34, 427-432 (1959). GORDON, S. A., and L. G. PALEG: Formation of auxin from tryptophane through the ac
tions of polyphenols. P!. Physio!. 36, 838-845 (1961). GORTNER, W. A., and M. J. KENT: The coenzyme requirement and enzyme inhibitors of
Pineapple indole acetic acid oxidase. J. Bio!. Chern. 233, 731-735 (1958). - Ferulic- and p-cumaric acids in Pineapple tissue as modifiers of Pineapple indole
acetic acid oxidase. Nature 181, 630-631 (1958 b). GREENWOOD, M. S., and M. H. M. GOLDSMITH: Polar transport and accumulation of indole-
3-acetic acid during root regeneration by Pinus lambertiana embryos. Plant a 95, 297-313 (1970).
HESS, C. E.: Rooting co factors - identification and function. Intern. Plant Prop. Soc. 15, 181-186 (1965).
- Phenolic compounds as stimulators of root initiation. Plant Phys., Supp!. 40, XLV (1965 b).
- Internal and external factors regulating root initiation. In: W. J. WHITTINGTON (Ed.), Root Growth, Proceedings of the fifteenth Easter School in Agricultural Science. University of Nottingham, 1968.
JANSSEN, M. G. H.: The influence of some phenolic cofactors on the pH-optimum of IAAoxydase from pea roots. Acta Bot. Neerl. 18,634-638 (1969).
-- Inhibitors of pea root indoleacetic acid oxidase. Acta Bot. Neerl. 19, 109-111 (1970). KAMINSKI, c.: Recherches sur les phenoloxydases dans les hypocotyles des Impatiens bal
samina L. Bul!. Acad, Roy. Belg. C!. Sci. 65, 154-168, 299-315 (1959). KEFELI, V. I.: Native Wachstumsinhibitoren, ihre physiologische Rolle und ihr Wirkungs
mechanismus. Wiss. Z. Uni Rostock 17, 383-389 (1968). KEFELI, V. I., and CH. SH. KADYROV: Natural growth inhibitors, their chemical and physio
logical properties. Ann. Rev. P!. Phys. 22, 185-195 (1971). KONINGS, H.: On the indoleacetic acid converting enzyme of the pea roots and its relation
to geotropism, straight growth and cell wall properties. Acta Bot. Neerl. 13, 566-622 (1964).
LEE, T. T., and F. SKOOG: Effect of hydroxybenzoic acid on IAA inactivation by tobacco callus extracts. Physio!. Plant. 18, 577-585 (1965).
LIB BERT, E.: Untersuchungen iiber die Physiologie der Adventivwurzelbildung. Flora 144, 121-150 (1956).
LIBBERT, E., A. DRAWERT, und R. SCHRODER: I. IAA-formation by sterile pea sections in vivo as influenced by IAA oxydase inhibitors and transaminase coenzymes. Physio!. Plant 22, 1217 (1969).
LIEBERMANN, M., and J. B. BIALE: Oxidative phophorylation by sweet potato mitochondria and its inhibition by polyphenols. P!. Phys. 31, 420-424 (1956).
MOYED, H. S., and V. WILLIAMSON: 3-methyleneoxindole of peas. P!. Phys. 42, 510-514 (1967).
NEISH, A. c.: Coumarins, phenylpropanes and lignins. In: J. BONNER and J. E. VARNER (Eds.), Plant Biochemistry. Academic Press, New York, London, 1965.
PILET, P. E.: Effect of chlorogenic acid on the auxin catabolism and the content of root tissues. Phytochem. 3, 617-621 (1964).
Z. Pjlanzenphysiol. Ed. 79. S. 109-120. 1976.
120 U. PINGEL
- Effect of p-hydroxybenzoic acid on growth, auxin content and auxin catabolism. Phytochern. 5, 77-82 (1966).
PILET, P. E., ]. P. ZRYD, et P. LAVANCHY: Role du pH dans l'analyse de I'activite auxinesoxydasique d'extraits bruts et purifies. Physio!. Veg. 5, 327-335 (1967).
PINGEL, u.: Der Einflu~ phenolischer Aktivatoren und Inhibitoren der IES-Oxydaseaktivitiit auf die Adventivbewurzelung bei Tradescantia albiflora. Dissertation Hamburg, 1974.
PRIDHAM, ]. B.: Low molecular weight phenols in higher plants. Ann. Rev. P!. Phys. 16, 13-36 (1965).
RABIN, R. S., and R. M. KLEIN: Chlorogenic acid as a competetive inhibitor of IAA-oxydase. Arch. Biochem. Biophys. 70, 11-15 (1957).
RIBEREAU-GAYON, P.: Plant Phenolics. University Reviews in Botany. Oliver and Boyd, Edinburgh, 1973.
RUGE, U.: Beeinflussung der Adventivwurzelbildung durch Stoffwechselgifte 1. Z. Pflanzenphys. 61, 430-446 (1969).
- Beeinflussung der Adventivwurzelbildung durch Stoffwechselgifte II. Z. Pflanzenphysio!. 65, 52-64 (1971).
RUNKOVA, L. V., E. K. LIs, M. TOMASZESKI, R. ANTOSZEWSKI: Function of phenolic substances in degradation system of indole-3-acetic acid in strawberries. Bio!. Plantarum 14, 71-81 (1972).
SACHER, ]. A.: Effect of inhibition on the kinetics of IAA oxidation. Americ. ]. Bot. 50, 116-122 (1963).
SHANTZ, E. M.: Chemistry and naturally occuring growth-regulating substances. Ann. Rev. P!. Phys. 17,409-438 (1966).
SOEKARJO, R.: On the formation of adventitious roots in cuttings of Coleus in relation to the effect of IAA on the epinastic curvature of isolated petioles. Acta. Bot. Neerl. 14, 373-399 (1965).
SOEKARJO, R., and M. G. H. JANSSEN: The liberation of inhibitors of IAA-oxydation activity out of Coleus internodes treated with KOH or H 2S04, 1969.
STAHL, E.: Diinnschichtchromatographie. Springer-Verlag, Berlin-Heidelberg-New York, 1967.
STENLID, G.: The effects of flavonoid compounds on oxidative phosphorylation and on the enzymic destruction of indole acetic acid. Physio!. Plant. 16, 110-120, 1963.
TOMASZEWSKI, M., and K. V. THIMANN: Interactions of phenolic acids, metallic ions and chelating agents of auxin induced growth. Plant Phys. 41, 1443-1454 (1966).
TULI, V., and H. S. MOYED: The role of 3-methyleneoxindole in auxin action. ]. Bio!. Chern. 244,4916-4920 (1969).
VAN SUMERE, C. F., G. WOLF, H. TEUCHY, and]. KINT: A new thin-layer method for phenolic substances and coumarins. ]. Chromat. 20, 48-60 (1965).
VARGA, M., and E. KavEs: Effect of phenolic compounds on the activity of indole-3-acetic acid oxidase. Naturwissenschaft 49, 55-56 (1962).
WEISER, C. ]., and L. T. BLANEY: The nature of boron stimulation to root initiation and development in beans. American Soc. Hort, Sc. 90, 191-200 (1967).
Dr. UWE PINGEL, Institut flir Angewandte Botanik der Universitiit, Marseiller Stra~e 7, D-2000 Hamburg 36.