24
Evolutionäre Verfahren 1 : Yeast Two Hybrid System Biochemiepraktikum II WS 2010/2011 18. Januar 2012 08. Februar 2012 AG Bernd Groner Laura Mack Axel Weber

Evolutionäre Verfahren 1 : Yeast Two Hybrid System
Biochemiepraktikum II WS 2010/2011
18. Januar 2012 08. Februar 2012
AG Bernd Groner Laura Mack Axel Weber
sd
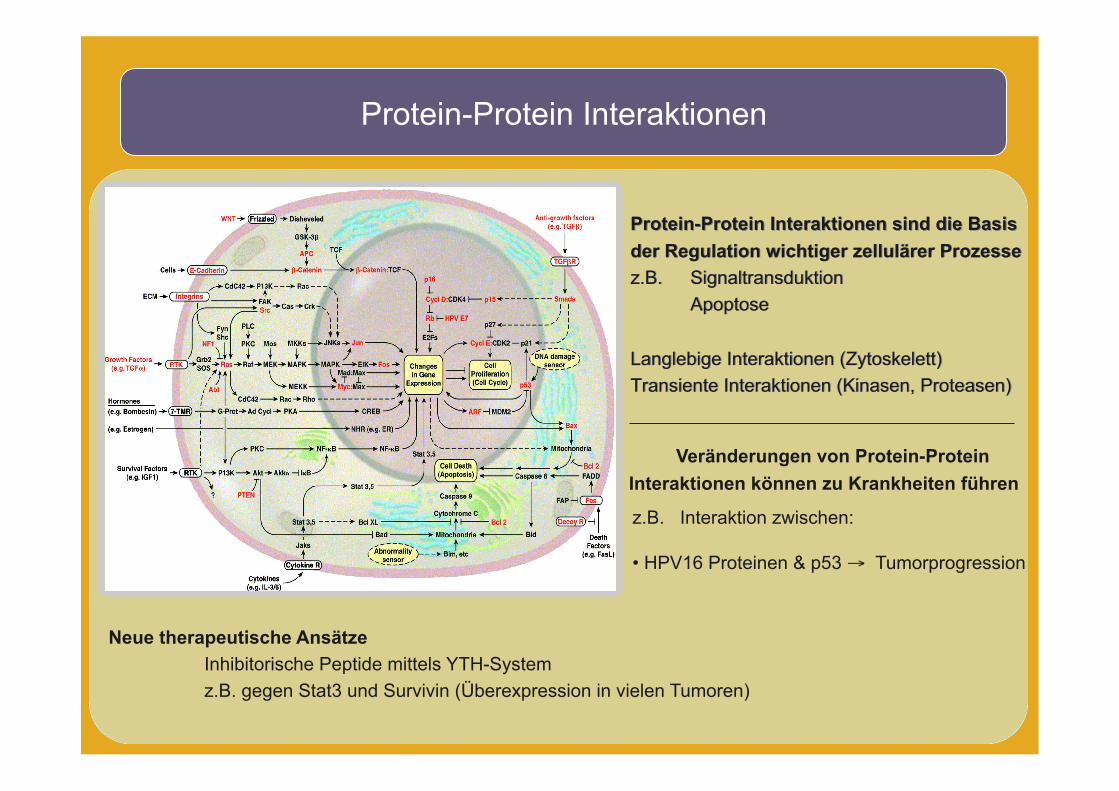
Protein-Protein Interaktionen
Protein-Protein Interaktionen sind die Basis der Regulation wichtiger zellulärer Prozesse z.B. Signaltransduktion Apoptose
Langlebige Interaktionen (Zytoskelett) Transiente Interaktionen (Kinasen, Proteasen)
Neue therapeutische Ansätze Inhibitorische Peptide mittels YTH-System z.B. gegen Stat3 und Survivin (Überexpression in vielen Tumoren)
Veränderungen von Protein-Protein Interaktionen können zu Krankheiten führen
z.B. Interaktion zwischen:
• HPV16 Proteinen & p53 → Tumorprogression

Analyse von Protein-Protein Interaktionen
Identifizierung => Charakterisierung => Manipulation
In vitro Methoden:
(Co-)Immunpräzipitation zwei interagierende Proteine im Lysat Affinitätschromatographie potentieller Interaktionspartner ist immobilisiert Cross-linking chemische Verknüpfung von Partnern Protein-probing markiertes Protein beweist eine Bindung Phage-display Suche nach Partner in einer Bibliothek
In vivo Methoden:
Fluorescence resonance energy transfer (FRET) Hefe-Zwei-Hybrid
Reverse-Zwei-Hybrid-System Hefe-Tribrid-System (Dual-Bait-System)
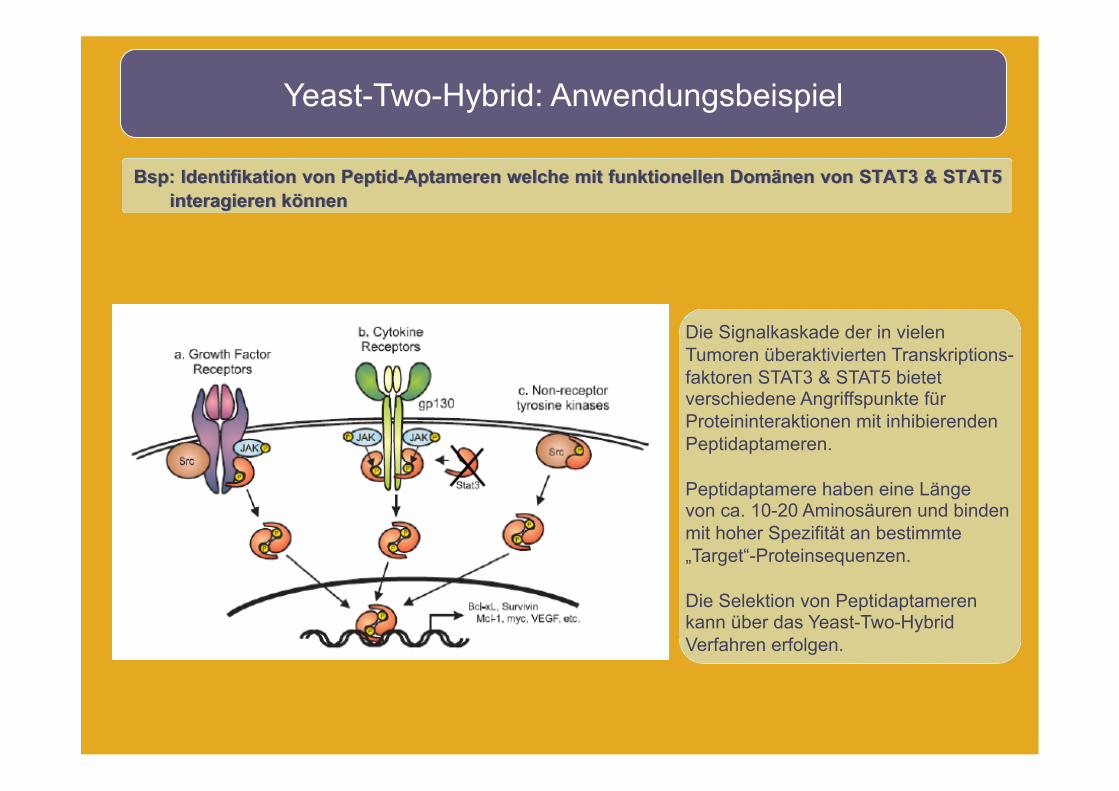
Yeast-Two-Hybrid: Anwendungsbeispiel
Bsp: Identifikation von Peptid-Aptameren welche mit funktionellen Domänen von STAT3 & STAT5 interagieren können
Die Signalkaskade der in vielen Tumoren überaktivierten Transkriptions- faktoren STAT3 & STAT5 bietet verschiedene Angriffspunkte für Proteininteraktionen mit inhibierenden Peptidaptameren.
Peptidaptamere haben eine Länge von ca. 10-20 Aminosäuren und binden mit hoher Spezifität an bestimmte „Target“-Proteinsequenzen.
Die Selektion von Peptidaptameren kann über das Yeast-Two-Hybrid Verfahren erfolgen.
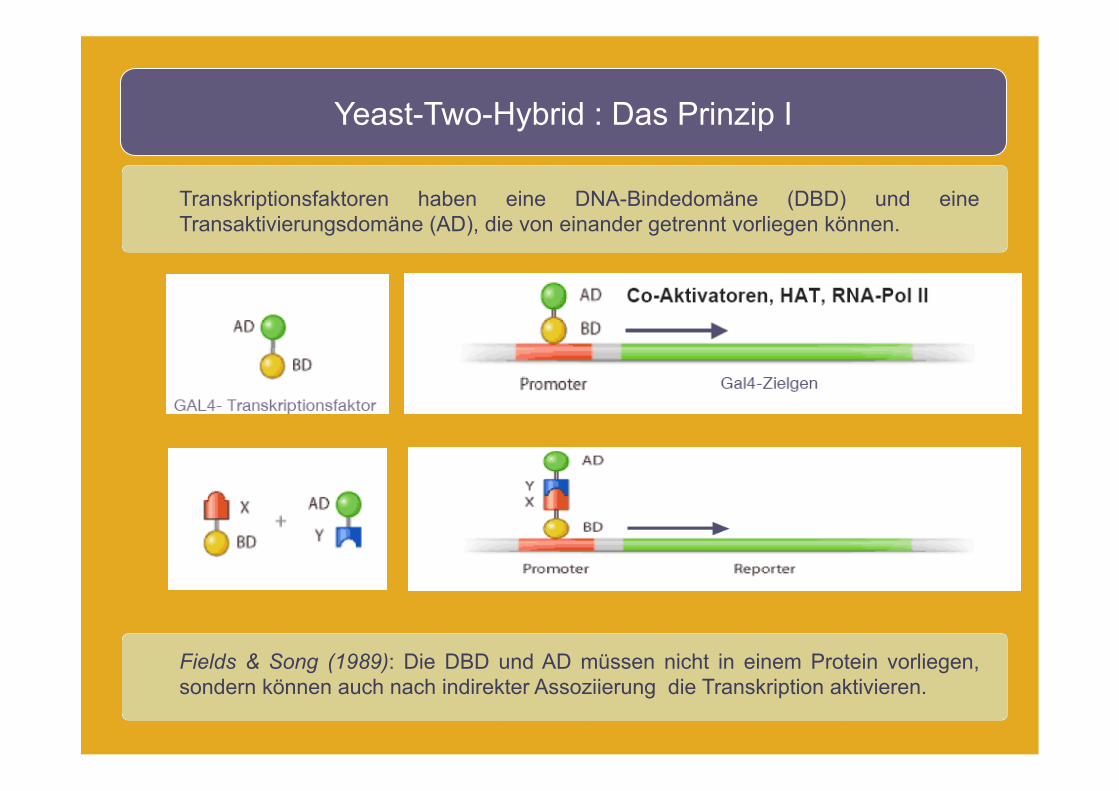
Yeast-Two-Hybrid : Das Prinzip I
Transkriptionsfaktoren haben eine DNA-Bindedomäne (DBD) und eine Transaktivierungsdomäne (AD), die von einander getrennt vorliegen können.
Fields & Song (1989): Die DBD und AD müssen nicht in einem Protein vorliegen, sondern können auch nach indirekter Assoziierung die Transkription aktivieren.
Ade2 His3
Gal4 RE
Protein 1 Gal4 DBD
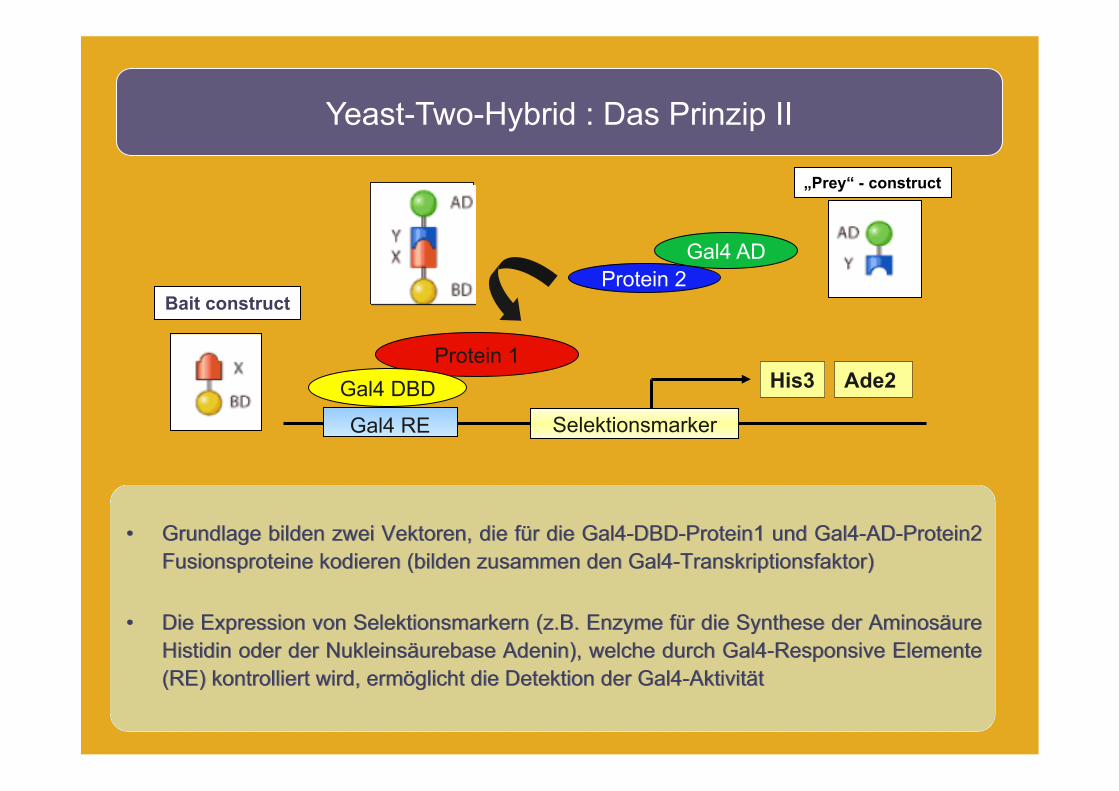
• Grundlage bilden zwei Vektoren, die für die Gal4-DBD-Protein1 und Gal4-AD-Protein2 Fusionsproteine kodieren (bilden zusammen den Gal4-Transkriptionsfaktor)
• Die Expression von Selektionsmarkern (z.B. Enzyme für die Synthese der Aminosäure Histidin oder der Nukleinsäurebase Adenin), welche durch Gal4-Responsive Elemente (RE) kontrolliert wird, ermöglicht die Detektion der Gal4-Aktivität
Yeast-Two-Hybrid : Das Prinzip II
Gal4 AD Protein 2
Selektionsmarker
„Prey“ - construct
Bait construct

Yeast-Two-Hybrid : Hefe
Genotyp:
MATa, his3-200, trp1-901, leu2-3,112, ura3-52, gal4, gal80, ade-2, LYS2::(GAL1UAS)-HIS3, LEU2::(GAL2UAS)-ADE2, URA3::(GAL1UAS)-LacZ
MATa oder α (mating types)
Reportergene:
His-3: IGP-dehydratase, ist an der Biosynthese von Histidin beteiligt
Ade-2: Phosphoribosylamino-imidazole-carboxylase ist an der Biosynthese von Adenin beteiligt
Ura-3: Orotidine-5’-phosphat decarboxylase, die an der Biosynthese der Pyrimidinbase Uracil beteiligt ist
LacZ: Kann verwendet werden, um die Interaktion im ß-Gal Test nachzuweisen
Deletion Modifikation
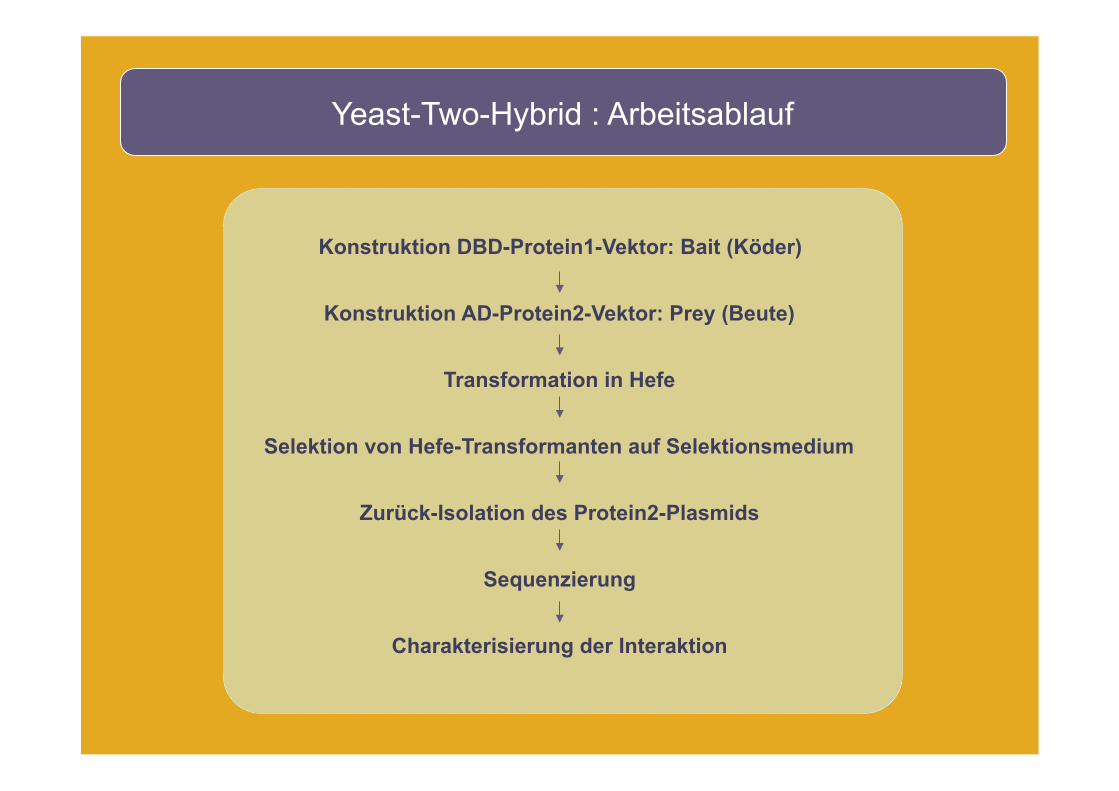
Yeast-Two-Hybrid : Arbeitsablauf
Konstruktion DBD-Protein1-Vektor: Bait (Köder)
Konstruktion AD-Protein2-Vektor: Prey (Beute)
Transformation in Hefe
Selektion von Hefe-Transformanten auf Selektionsmedium
Zurück-Isolation des Protein2-Plasmids
Sequenzierung
Charakterisierung der Interaktion
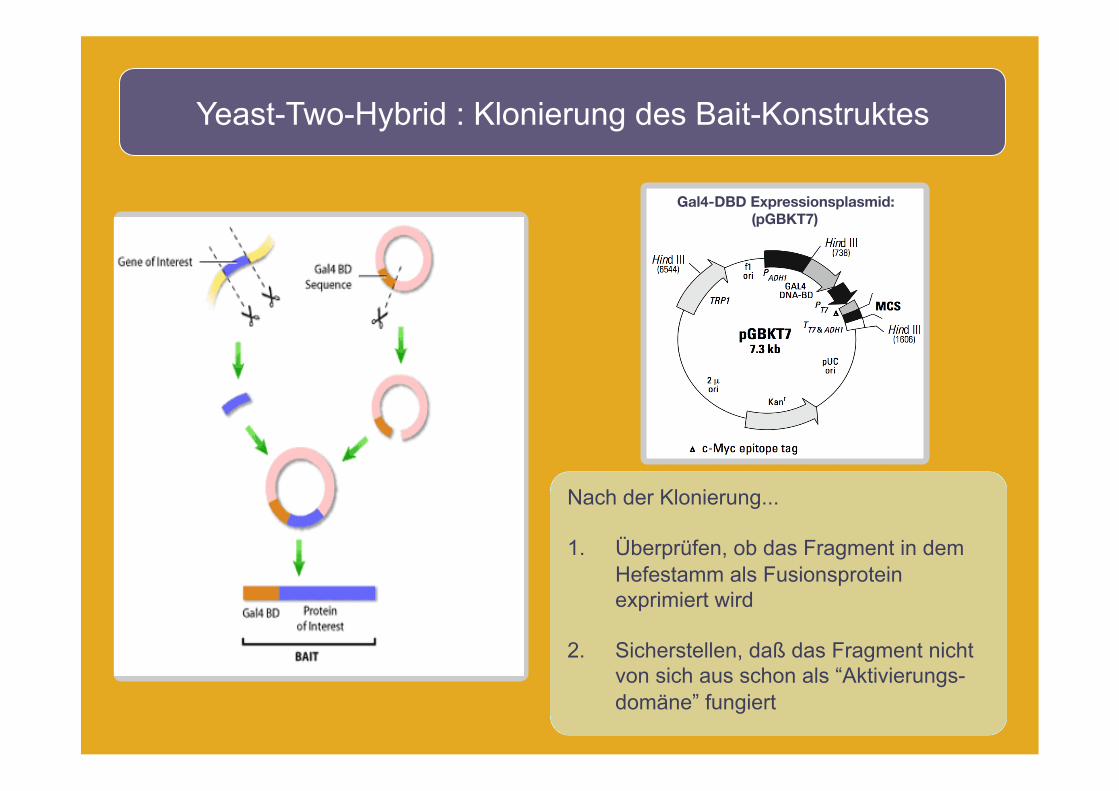
Yeast-Two-Hybrid : Klonierung des Bait-Konstruktes
Nach der Klonierung...
1. Überprüfen, ob das Fragment in dem Hefestamm als Fusionsprotein exprimiert wird
2. Sicherstellen, daß das Fragment nicht von sich aus schon als “Aktivierungs-domäne” fungiert
Gal4-DBD Expressionsplasmid: (pGBKT7)
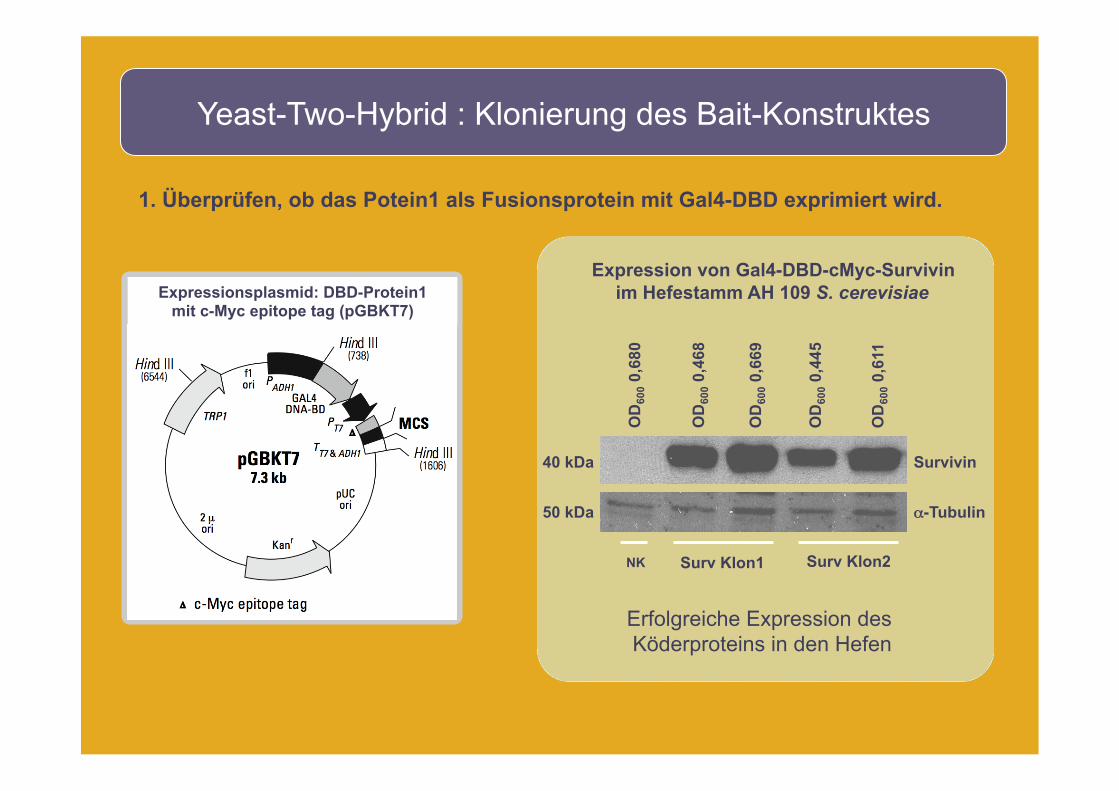
Yeast-Two-Hybrid : Klonierung des Bait-Konstruktes
1. Überprüfen, ob das Potein1 als Fusionsprotein mit Gal4-DBD exprimiert wird.
Survivin
α-Tubulin
OD
600 0
,468
OD
600 0
,669
OD
600 0
,445
OD
600 0
,611
OD
600 0
,680
Surv Klon1 Surv Klon2
40 kDa
50 kDa
Erfolgreiche Expression des Köderproteins in den Hefen
Expression von Gal4-DBD-cMyc-Survivin im Hefestamm AH 109 S. cerevisiae
NK
Expressionsplasmid: DBD-Protein1 mit c-Myc epitope tag (pGBKT7)

Yeast-Two-Hybrid : Konstruktion der “Prey”-Expressionsvektoren
Prey: 1. Ein zweites bekanntes Protein, oder ein Fragment dieses Proteins
2. cDNA- bzw. Peptid-Bank
3. Peptid-Bank integriert in einem Gerüstprotein
Gal4-AD cDNA
Gal4-AD Protein2
Gal4-AD Peptid Gerüst
Subdomäne *
Peptid
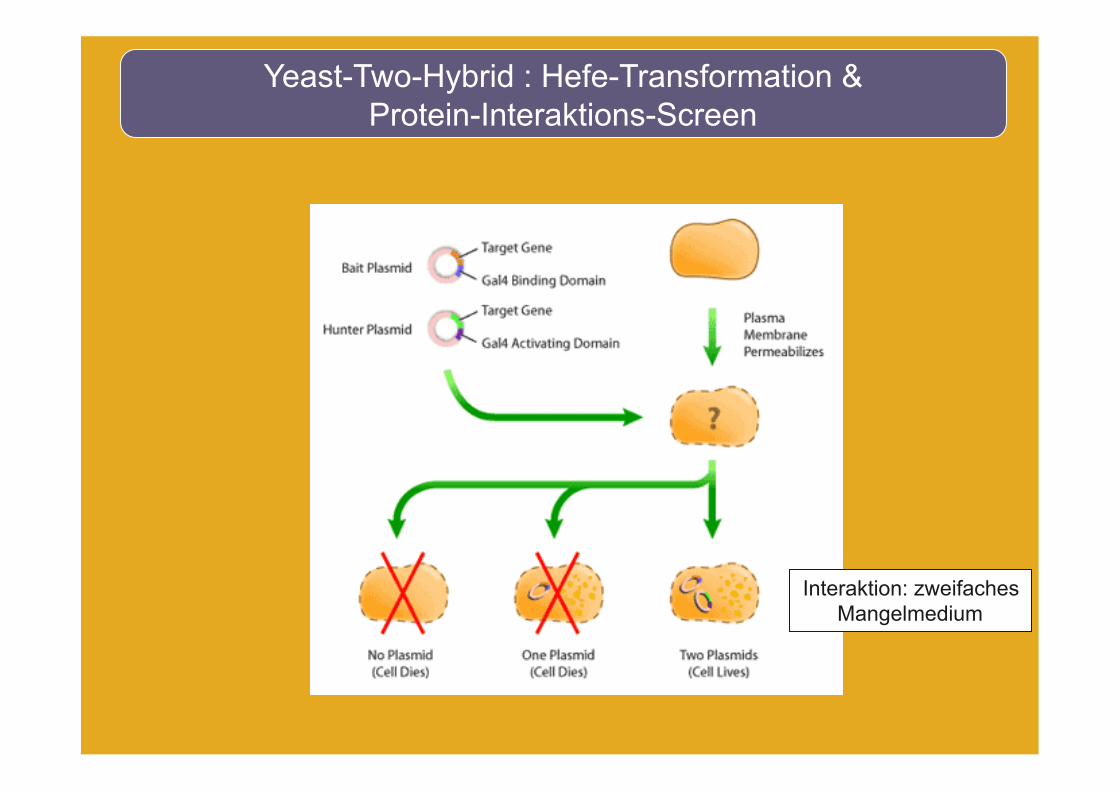
Yeast-Two-Hybrid : Hefe-Transformation & Protein-Interaktions-Screen
Interaktion: zweifaches Mangelmedium
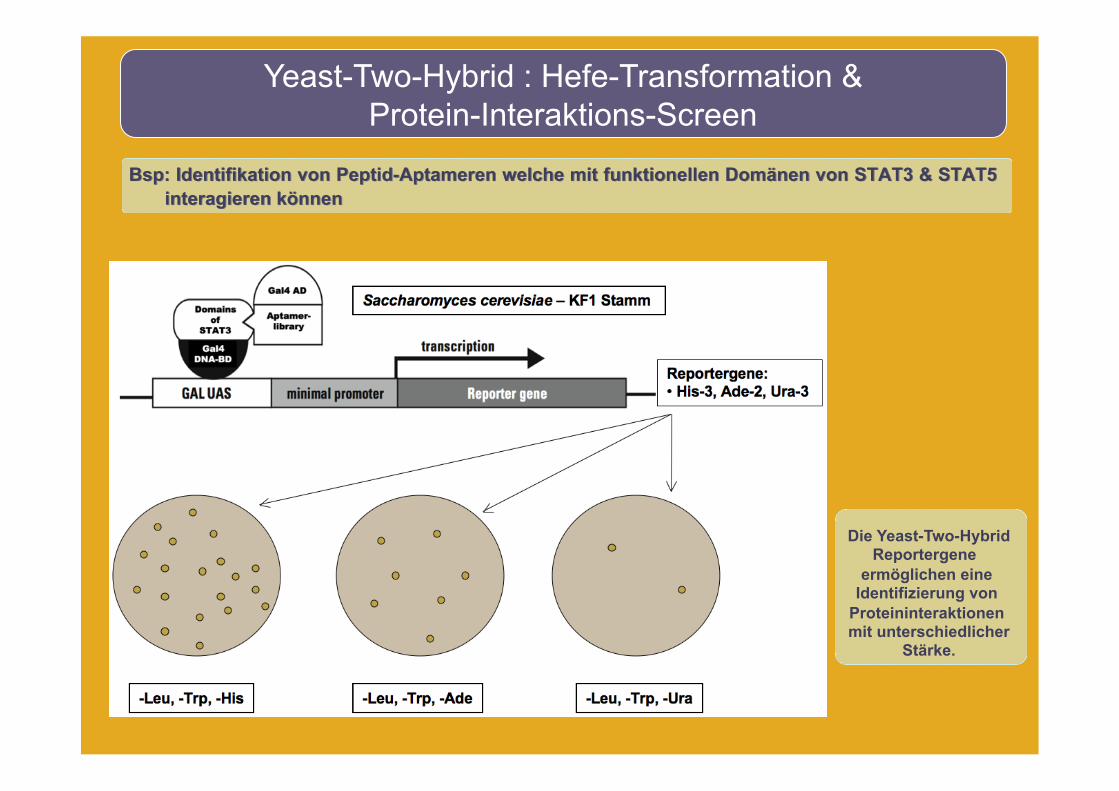
Yeast-Two-Hybrid : Hefe-Transformation & Protein-Interaktions-Screen
Bsp: Identifikation von Peptid-Aptameren welche mit funktionellen Domänen von STAT3 & STAT5 interagieren können
Die Yeast-Two-Hybrid Reportergene
ermöglichen eine Identifizierung von
Proteininteraktionen mit unterschiedlicher
Stärke.
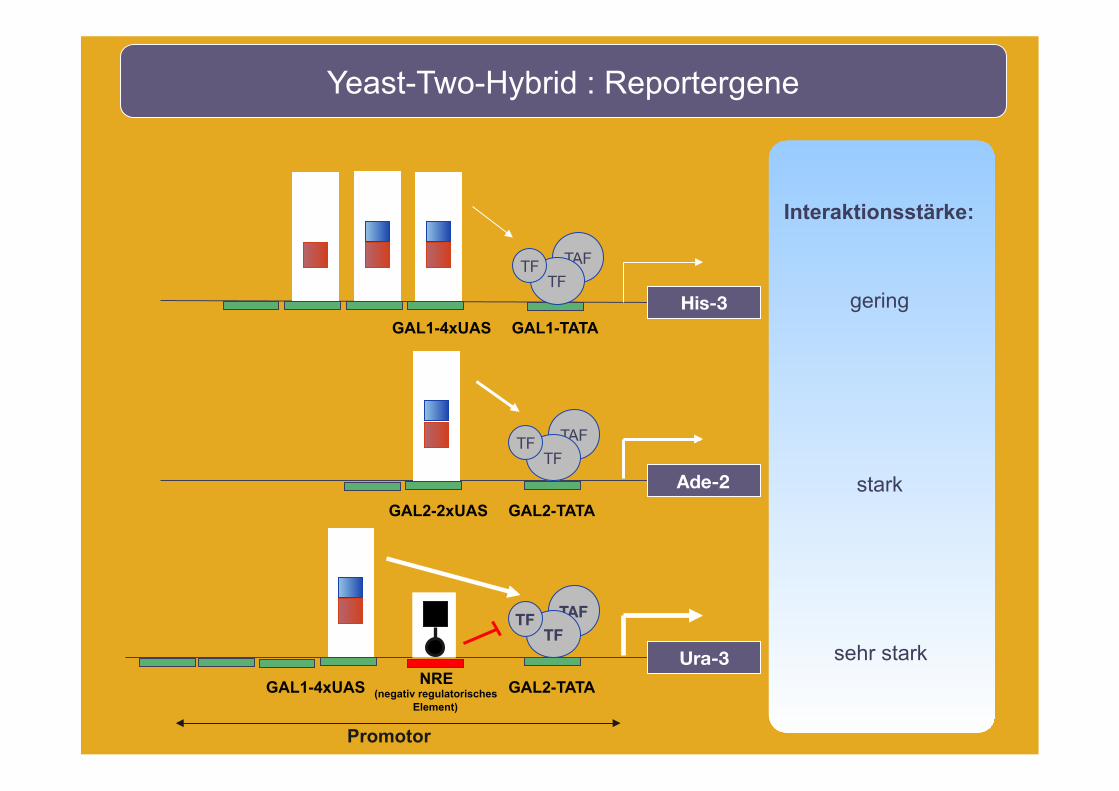
TAF
Yeast-Two-Hybrid : Reportergene
Interaktionsstärke:
His-3
Ade-2
GAL1-4xUAS GAL1-TATA
GAL2-2xUAS GAL2-TATA
gering
stark
TF TF
TAF
TF TF
Promotor
Ura-3
GAL1-4xUAS GAL2-TATA
TAF TF
TF
NRE (negativ regulatorisches
Element)
sehr stark
sd

Yeast-Two-Hybrid : Reportergene
-‐LT -‐LTH -‐LTA -‐LTU
GBD-DBD (STAT5) + GAD-S5-DBD-PA
100 10-1 10-2 10-3 100 10-1 10-2 10-3 100 10-1 10-2 10-3 100 10-1 10-2 10-3
PK (GBD-p53 + GAD-LT)
NK (GBD-LamC + GAD-LT)
GBD + GAD-S5-DBD-PA
GBD-SH2(STAT3) + GAD-r-S3-PA
GBD-SH2 (STAT5) + GAD-S5-DBD-PA
GBD-DBD (STAT3) + GAD-S5-DBD-PA
GBD-SH2 (STAT3) + GAD-S5-DBD-PA
bait + prey
Bsp: Identifikation von Peptid-Aptameren welche mit funktionellen Domänen von STAT3 & STAT5 interagieren können
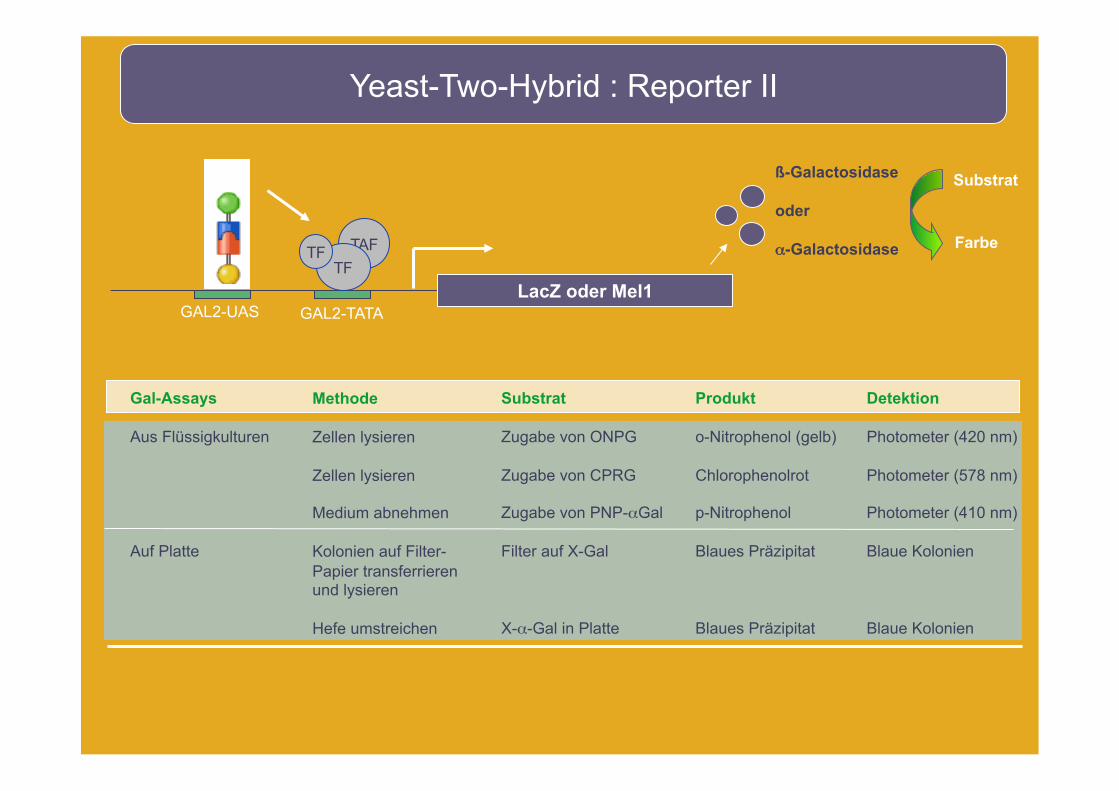
Yeast-Two-Hybrid : Reporter II
LacZ oder Mel1 GAL2-UAS GAL2-TATA
TAF
TF TF
Gal-Assays
Aus Flüssigkulturen
Auf Platte
Methode
Zellen lysieren
Zellen lysieren
Medium abnehmen
Kolonien auf Filter- Papier transferrieren und lysieren
Hefe umstreichen
Substrat
Zugabe von ONPG
Zugabe von CPRG
Zugabe von PNP-αGal
Filter auf X-Gal
X-α-Gal in Platte
Produkt
o-Nitrophenol (gelb)
Chlorophenolrot
p-Nitrophenol
Blaues Präzipitat
Blaues Präzipitat
Detektion
Photometer (420 nm)
Photometer (578 nm)
Photometer (410 nm)
Blaue Kolonien
Blaue Kolonien
ß-Galactosidase
oder
α-Galactosidase
Substrat
Farbe
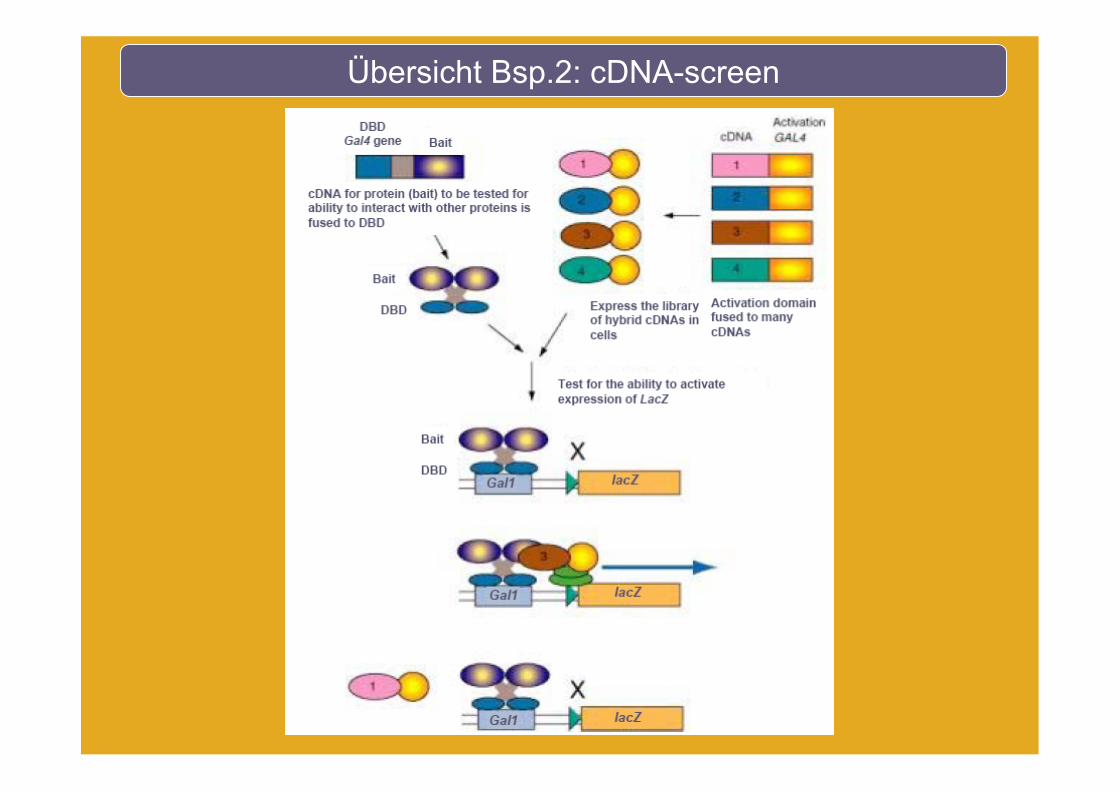
Übersicht Bsp.2: cDNA-screen
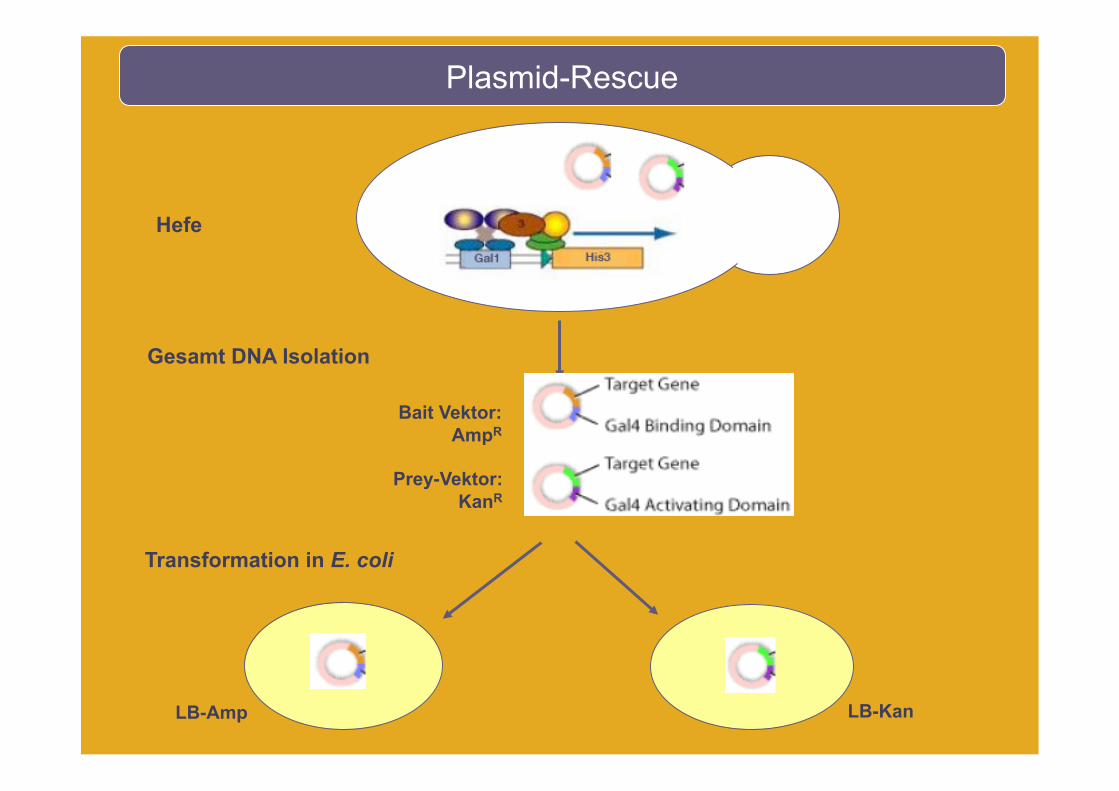
Plasmid-Rescue
Gesamt DNA Isolation
Transformation in E. coli
Bait Vektor: AmpR
Prey-Vektor: KanR
LB-Kan
Hefe
LB-Amp
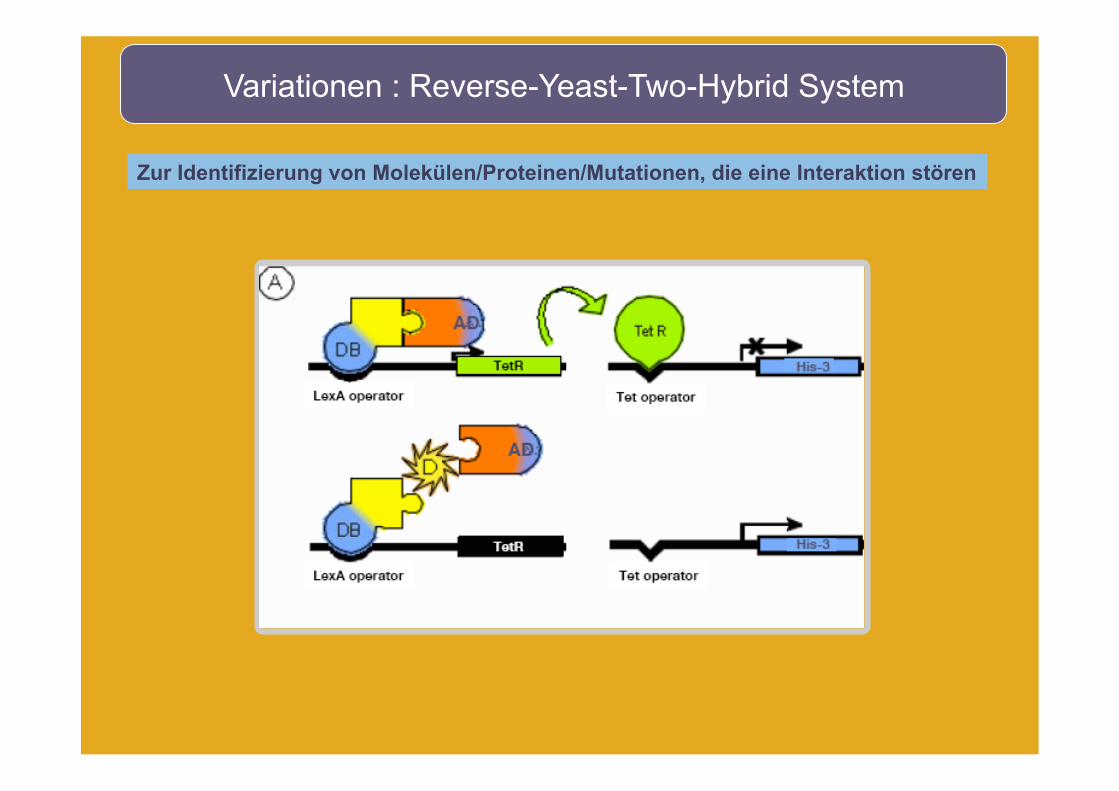
Variationen : Reverse-Yeast-Two-Hybrid System
Zur Identifizierung von Molekülen/Proteinen/Mutationen, die eine Interaktion stören
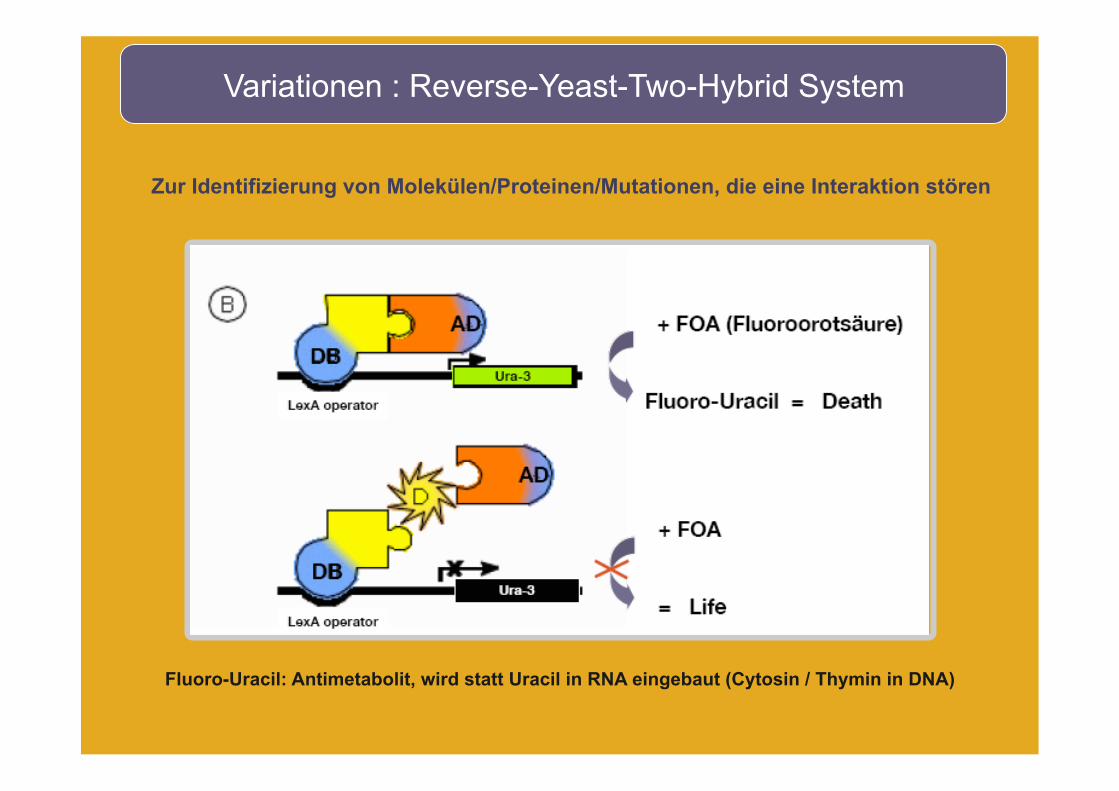
Variationen : Reverse-Yeast-Two-Hybrid System
Zur Identifizierung von Molekülen/Proteinen/Mutationen, die eine Interaktion stören
Fluoro-Uracil: Antimetabolit, wird statt Uracil in RNA eingebaut (Cytosin / Thymin in DNA)
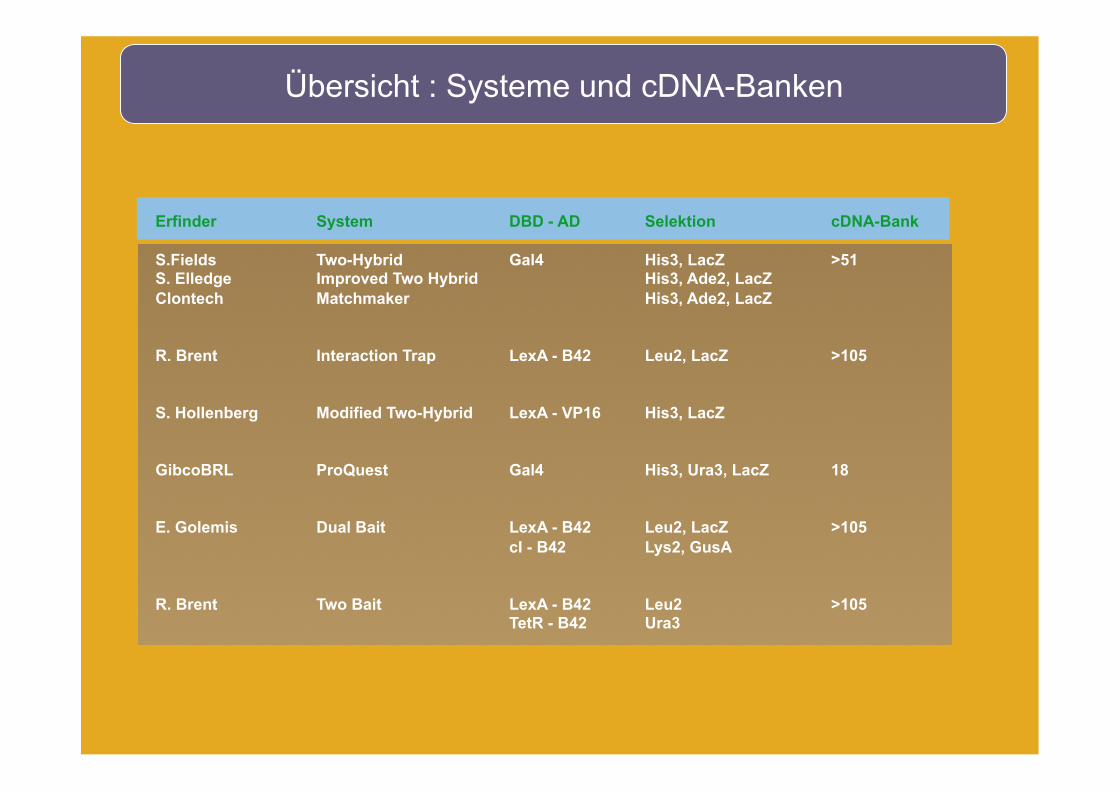
Übersicht : Systeme und cDNA-Banken
Erfinder
S.Fields S. Elledge Clontech
R. Brent
S. Hollenberg
GibcoBRL
E. Golemis
R. Brent
System
Two-Hybrid Improved Two Hybrid Matchmaker
Interaction Trap
Modified Two-Hybrid
ProQuest
Dual Bait
Two Bait
DBD - AD
Gal4
LexA - B42
LexA - VP16
Gal4
LexA - B42 cI - B42
LexA - B42 TetR - B42
Selektion
His3, LacZ His3, Ade2, LacZ His3, Ade2, LacZ
Leu2, LacZ
His3, LacZ
His3, Ura3, LacZ
Leu2, LacZ Lys2, GusA
Leu2 Ura3
cDNA-Bank
>51
>105
18
>105
>105

Vorteile des Hefe-Zwei-Hybrid Systems
1. In vivo
2. Es ist relativ einfach hiermit anzufangen und benötigt nicht viel Aufwand. Für biochemische Methoden wird oft aufgereinigtes Protein in sehr guter Qualität benötigt. Hier wird höchstens cDNA benötigt.
3. Hiermit können auch die schwachen Interaktionen noch detektiert werden, da mehrfache Promoterelemente vor dem Reportergen geschaltet sind, was zur Amplifikation des Signals führt.
4. Es können ganz einfache semi-quantitative Tests durchgeführt werden, um die Affinität interpretieren zu können.
5. Nach dem Screen kann sofort die DNA des Interaktionspartners isoliert werden.

Zu beachten : Potentielle Nachteile
1. Das Target-Protein darf keine eigene Aktivierungsdomäne besitzen. 2. Die Konformation des Target-Proteins kann im Fusionsprotein geändert sein. Wird
nur eine Domäne verwendet, soll unbedingt die Bindung am gesamten Protein untersucht werden.
3. Wird nur eine bestimmte Domäne des Target-Proteins an die DBD fusioniert (weil das Target zu groß ist oder weil es sich um ein Membranprotein handelt) dann soll beachtet werden, dass so normal nicht zugängliche Bindungsstellen vielleicht jetzt an der Oberfläche liegen.
4. Ein Protein, von dem bekannt ist, dass es das Target-Protein bindet, sollte als Positiv-Kontrolle verwendet werden (bei Interaktion ist die Faltung der Domäne korrekt).
5. Manche post-translationale Modifikationen finden nicht statt. 6. Die Fusionsproteine müssen in den Kern gelangen. Sind stärkere
Translokationssignale im Fusionsprotein vorhanden (z.B. Sekretion)? 7. Es kann nicht ausgeschlossen werden, dass ein drittes Protein für die Interaktion
benötigt wird. 8. Hefe wird als Wirt benutzt.
Nachteil: Ist die Faltung korrekt und das Protein in Hefen stabil? Vorteil: Gleicht mehr der Situation in höheren Eukaryoten als bei in vitro
Experimenten oder Systemen mit Bakterien als Wirt

Vielen Dank für Eure Aufmerksamkeit !

















