Page 1
Aufnahme und Verteilung von Lindan in Tomatenpflanzen
Abschlussarbeit Postgradualstudium
Toxikologie und Umweltschutz
Universität Leipzig
Dipl. – Geoökologin Nadine Zeiner
Leipzig, 12.12.2007
Page 2
__________ Inhaltsverzeichnis
2
1 Einleitung ......................................................................................................................... 3
2 HCH in der Umwelt .......................................................................................................... 4
2.1 Eigenschaften und Verwendung ............................................................................. 4
2.2 Verhalten von HCH in der Umwelt ........................................................................... 7
2.3 Aufnahme von organischen Umweltchemikalien in Pflanzen .................................. 8
2.4 Aufnahme und Metabolismus von HCH in Pflanzen .............................................. 10
3.1 Aufnahme, Verteilung, Metabolismus und Ausscheidung (Elimination) von HCH durch den Menschen ..................................................................................................... 13
3.2 Akute Toxizität ....................................................................................................... 15
3.3 Chronische Toxizität ............................................................................................... 17
3.4 Spezielle Toxikologie ............................................................................................. 18
4.1 Versuchsdurchführung ........................................................................................... 20
4.2 Analytik ................................................................................................................... 21
4.2.1 Probenvorbereitung ......................................................................................... 21
4.2.2 Analytik ............................................................................................................ 26
5 Ergebnisse ................................................................................................................... 27
5.1 Rückstandsanalytik in der Nährlösung und im Boden ............................................ 27
5.2 Lindankonzentrationen in den Versuchspflanzen ................................................... 27
5.3 Lindankonzentrationen in Früchten ........................................................................ 31
6 Zusammenfassung ......................................................................................................... 32
Literaturverzeichnis ........................................................................................................... 33
Tabellenverzeichnis .......................................................................................................... 37
Abbildungsverzeichnis ...................................................................................................... 37
Abkürzungsverzeichnis ..................................................................................................... 38
Page 3
1 Einleitung
3
1 Einleitung
Hexachlorcyclohexan (HCH) gehört zur Gruppe der Organochlorpestizide.
Technisches HCH wurde seit den 1940er Jahren als Insektizid u. a. in der
Landwirtschaft verwendet. Dieses Technische HCH besteht aus 8 verschiedenen
Isomeren (Hauptisomere: α-, β-, γ- und δ-HCH), von denen nur γ-HCH eine
insektizide Wirkung aufweist. Unter dem Namen Lindan kam ein Produkt auf den
Markt, welches zu mindestens 99% aus γ-HCH besteht.
Der Einsatz von HCH bzw. Lindan in der Landwirtschaft ist seit 1977 in der BRD und
seit 2003 europaweit verboten. In der DDR wurde Lindan noch bis Ende der 1980er
Jahre genutzt. Heute kommt Lindan noch als Biozid in der Human- und
Veterinärmedizin zur Anwendung. Außerdem wird im außereuropäischen Ausland
technisches HCH und Lindan weiterhin genutzt.
Obwohl Lindan heute in Europa nicht mehr großflächig eingesetzt wird, findet man es
aufgrund seiner Persistenz und Mobilität in den Umweltkompartimenten (Boden,
Wasser, Luft) sowie an ausgewählten Standorten in pflanzlichen Nahrungsmitteln und
im menschlichen Gewebe. Somit müssen die HCH – Isomere zu den ubiquitären
Umweltschadstoffen gezählt werden. Aufgrund der Persistenz, der Mobilität in der
Umwelt, der Fähigkeit zur Bioakkumulation sowie der potentiellen Toxizität und
Kanzerogenität sind die HCH-Isomere als kritische Umweltschadstoffe anzusehen.
Da HCH, aufgrund seiner Lipophilie, dazu neigt, sich in der Nahrungskette
anzureichern und die vorrangige Exposition des Menschen über kontaminierte
Lebensmittel führt, ist es wichtig, mögliche Expositionsquellen zu identifizieren. In
verschiedenen Untersuchungen [u. a. Li, 2002; Gao, 2005; Gonzalez, 2003; Verma
und Pillai, 1991] wurde bereits gezeigt, dass Lindan häufig in Nutzpflanzen und
Gemüseproben gefunden wird.
In der vorliegenden Arbeit wird zunächst auf die toxikologische Relevanz der HCH-
Isomere, besonders von γ-HCH, eingegangen. Im praktischen Teil werden Tomaten-
pflanzen mit Lindan exponiert und der Transfer der Substanz in die Pflanze wird
untersucht. Um die Verteilung innerhalb der Pflanze nachzuvollziehen, werden
Rückstandbestimmungen in Boden und Nährlösung sowie Analysen der einzelnen
Pflanzenteile durchgeführt. Um ein potentielles Risiko des Verbrauchers durch den
Verzehr der Früchte zu beurteilen, werden die Ergebnisse mit Grenz- und Richtwerten
verglichen.
Page 4
2 HCH in der Umwelt
4
2 HCH in der Umwelt 2.1 Eigenschaften und Verwendung
1,2,3,4,5,6-Hexachlorcyclohexan (HCH) ist ein monocyclischer chlorierter
Kohlenwasserstoff. Der Cyclohexanring ist nicht planar, sondern liegt, wie aus Abb. 1
ersichtlich, in Sesselform (trans-Form) vor. Diese Struktur ist energieärmer als die
stereoisomere Wannenform und daher stabiler als diese.
Abb. 1: Darstellung des HCH in der Sesselform [Römpp, 1983]
Die Substituenten können entweder eine axiale (a) oder eine äquatoriale (e) Lage zur
Ringebene einnehmen. Je nach Stellung der Chloratome entstehen so 8 Isomere, die in
Abb. 2 dargestellt sind. Die HCH-Isomere unterscheiden sich durch ihre kristalline und
räumliche Struktur. Von diesen Isomeren sind lediglich 5 in nennenswerten Mengen im
technischen HCH enthalten [LfU, 1993; Marquardt & Schäfer, 1994]. HCH kommt nicht
als natürliche Substanz vor. Technisches Hexachlorcyclohexan wird durch Chlorierung
von Benzol unter UV-Licht gewonnen. Es enthält etwa 65 – 70 % α-HCH, 10 % β-HCH,
15 % γ- HCH, 7 % δ- HCH sowie weitere HCH-Isomere in geringerer Konzentration
[Marquardt & Schäfer, 1994]. Als Lindan bezeichnet man das Produkt, das zu mindestens
99% aus γ-HCH besteht [UBA, 2006].
Page 5
2 HCH in der Umwelt
5
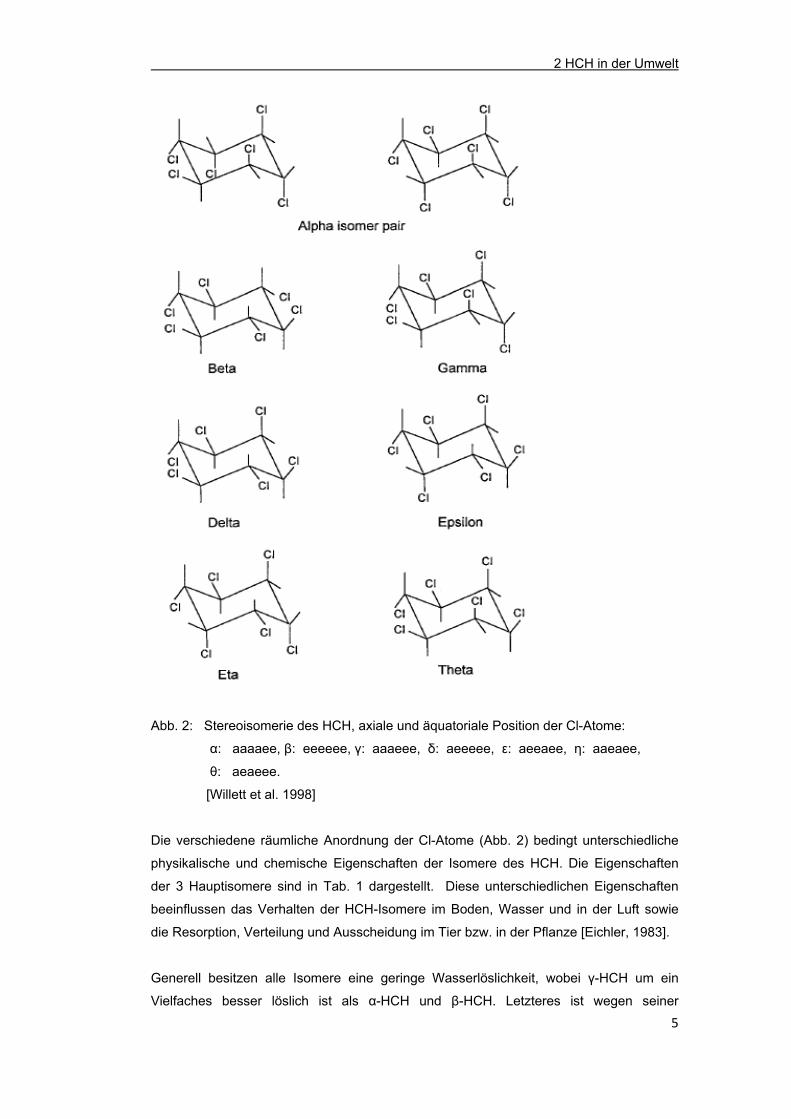
Abb. 2: Stereoisomerie des HCH, axiale und äquatoriale Position der Cl-Atome:
α: aaaaee, β: eeeeee, γ: aaaeee, δ: aeeeee, ε: aeeaee, η: aaeaee,
θ: aeaeee.
[Willett et al. 1998]
Die verschiedene räumliche Anordnung der Cl-Atome (Abb. 2) bedingt unterschiedliche
physikalische und chemische Eigenschaften der Isomere des HCH. Die Eigenschaften
der 3 Hauptisomere sind in Tab. 1 dargestellt. Diese unterschiedlichen Eigenschaften
beeinflussen das Verhalten der HCH-Isomere im Boden, Wasser und in der Luft sowie
die Resorption, Verteilung und Ausscheidung im Tier bzw. in der Pflanze [Eichler, 1983].
Generell besitzen alle Isomere eine geringe Wasserlöslichkeit, wobei γ-HCH um ein
Vielfaches besser löslich ist als α-HCH und β-HCH. Letzteres ist wegen seiner
Page 6
2 HCH in der Umwelt
6
symmetrischen Struktur völlig unpolar, was zu einer stärkeren Anreicherung in Fett im
Vergleich zu den anderen Isomeren führt. α-HCH und γ-HCH zeigen dagegen höhere
Werte für das Dipolmoment als β-HCH. Auch im Hinblick auf den Dampfdruck und den
Schmelzpunkt verhält sich β-HCH abweichend von den anderen Isomeren [Eichler,
1983].
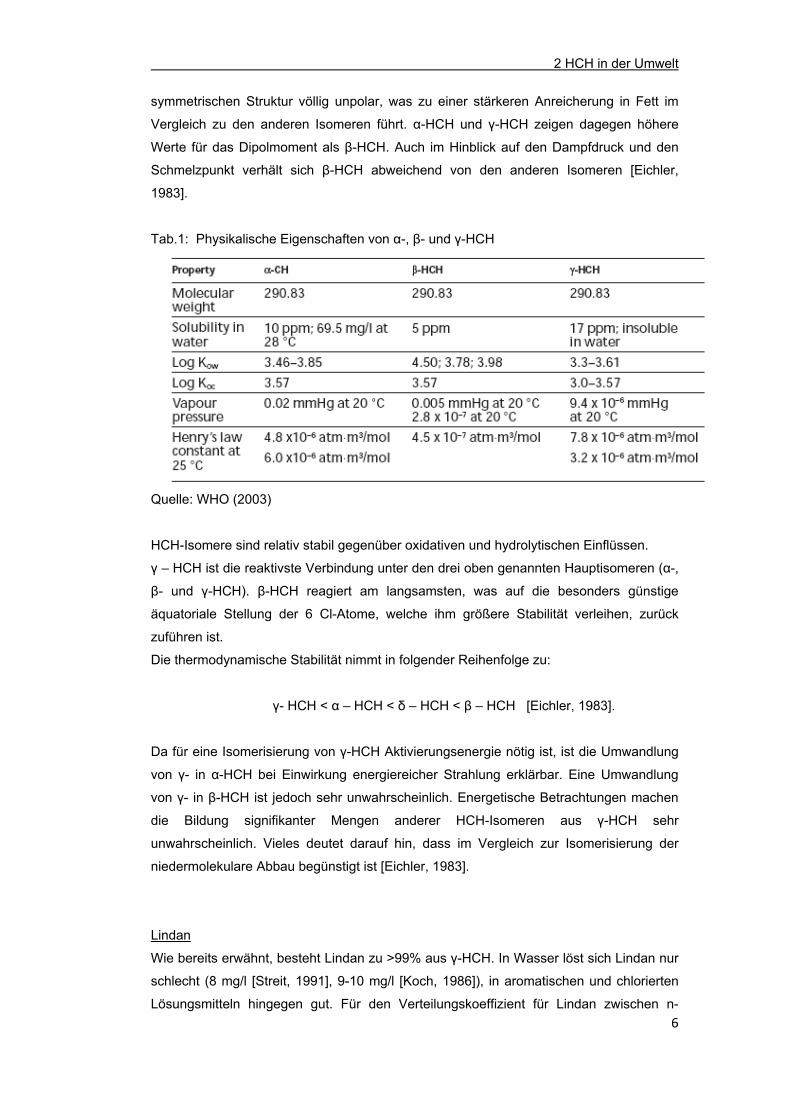
Tab.1: Physikalische Eigenschaften von α-, β- und γ-HCH
Quelle: WHO (2003)
HCH-Isomere sind relativ stabil gegenüber oxidativen und hydrolytischen Einflüssen.
γ – HCH ist die reaktivste Verbindung unter den drei oben genannten Hauptisomeren (α-,
β- und γ-HCH). β-HCH reagiert am langsamsten, was auf die besonders günstige
äquatoriale Stellung der 6 Cl-Atome, welche ihm größere Stabilität verleihen, zurück
zuführen ist.
Die thermodynamische Stabilität nimmt in folgender Reihenfolge zu:
γ- HCH < α – HCH < δ – HCH < β – HCH [Eichler, 1983].
Da für eine Isomerisierung von γ-HCH Aktivierungsenergie nötig ist, ist die Umwandlung
von γ- in α-HCH bei Einwirkung energiereicher Strahlung erklärbar. Eine Umwandlung
von γ- in β-HCH ist jedoch sehr unwahrscheinlich. Energetische Betrachtungen machen
die Bildung signifikanter Mengen anderer HCH-Isomeren aus γ-HCH sehr
unwahrscheinlich. Vieles deutet darauf hin, dass im Vergleich zur Isomerisierung der
niedermolekulare Abbau begünstigt ist [Eichler, 1983].
Lindan
Wie bereits erwähnt, besteht Lindan zu >99% aus γ-HCH. In Wasser löst sich Lindan nur
schlecht (8 mg/l [Streit, 1991], 9-10 mg/l [Koch, 1986]), in aromatischen und chlorierten
Lösungsmitteln hingegen gut. Für den Verteilungskoeffizient für Lindan zwischen n-
Page 7
2 HCH in der Umwelt
7
Oktanol und Wasser (lg Kow) werden Werte zwischen 3,2 (22 °C) [LfU, 1993] und 3,43
[Streit, 1991] angegeben.
Von allen Isomeren weist nur das γ-HCH (Lindan) eine insektizide Wirkung auf
[Marquardt und Schäfer, 1994].
Lindan wirkt als Insektizid mit Kontakt-, Fraß- und Atemgiftwirkung vor allem gegen
Bodenschädlinge (in Form von Saatgutbehandlungsmitteln) und gegen
rindenbewohnende Forstschädlinge.
Es ist Bestandteil von Holzschutzmitteln und wird im außereuropäischen Bereich bei der
Bekämpfung von Parasiten an Nutztieren verwendet. Außerdem wird Lindan gegen
Vorratsschädlinge und Wanzen in Kakaoplantagen und gegen Schadkäfer im
Kaffeeanbau eingesetzt [Falbe & Regitz, 1997].
Bis in die späten sechziger Jahre kam das technische HCH in der Forstwirtschaft zur
Anwendung [Korte, 1983]. Inzwischen ist auch Lindan (γ-HCH) als Pflanzenschutzmittel
wegen der hohen Persistenz in Deutschland verboten. Der Einsatz als Biozid in der
Human- und Veterinärmedizin (z. B. in Anti-Läuse-Shampoos) ist erlaubt [Scheffer &
Schachtschabel, 2002]. Europaweit gilt seit 2003 ein Verbot aller noch verbliebenen
landwirtschaftlichen Anwendungen [UBA, 2006].
2.2 Verhalten von HCH in der Umwelt
Umweltkontaminationen durch HCH ergeben sich aus der früheren Verwendung HCH-
haltiger Produkte in der Land- und Forstwirtschaft sowie in der Veterinär- und
Humanmedizin. Außerdem kommt es durch deponierte Produktionsrückstände aus der
Lindanherstellung zu Kontaminationen der Umwelt [UBA, 2006]. Aufgrund seiner
Persistenz ist es heute immer noch in allen Umweltkompartimenten nachweisbar [Fabre
et al. 2005].
Das Verhalten von HCH in der Umwelt wird vor allem durch seine im Vergleich zu
anderen chlororganischen Insektiziden mittlere Wasserlöslichkeit und Flüchtigkeit sowie
der daraus resultierenden hohen Bio- und Geoakkumulationstendenz und Mobilität in und
zwischen den Umweltmedien Wasser, Boden und Luft beeinflusst [Koch, 1986]. Aufgrund
des unterschiedlichen Grades der Lipophilie der Isomere, ist das Potential zur
Anreicherung in biologischen Systemen beim β-HCH am stärksten ausgeprägt und nimmt
über das α-, γ- und δ-HCH ab [Marquardt & Schäfer, 1994; Braun et al. 1999].
In natürlichen Wässern ist HCH relativ stabil gegenüber physikalisch-chemischem Abbau
[Koch, 1986]. Photolyse und Hydrolyse scheinen langsam zu verlaufen [Streit, 1991]. Es
erfolgt eine relativ schnelle Sorption an partikuläre Stoffe bzw. Sedimente sowie eine
Page 8
2 HCH in der Umwelt
8
Bioakkumulation vor allem in Phyto- und Zooplankton. Metabolisiert wird die Verbindung
in Abhängigkeit von der biologischen Aktivität der Gewässer und Sedimente.
In Böden wird die Verbindung abhängig vom organischen Gehalt der Bodenmatrix
vorzugsweise in oberflächennahen Schichten adsorbiert. Aufgrund der Wasserlöslichkeit
sind jedoch Migrationen in grundwasserführende Schichten nicht auszuschließen [Koch,
1986]. Der Abbau der HCH-Isomere im Boden hängt von Bodenart, Bodentyp und
Umgebungsbedingungen, wie Bewässerung, Bewuchs und Bodenbearbeitung, ab. Dabei
spielen mikrobiologische sowie chemische Vorgänge eine Rolle. Auch Prozesse wie
Verflüchtigung und Photolyse sind relevant [LfU, 1993]. Trotz ihrer Toxizität (z. B.
Hemmung der Zellteilung durch α-HCH bei Bakterien) können die HCH-Isomere
biologisch metabolisiert werden. Aus Lindan können durch Biotransformation neben α-
HCH auch β- und δ-HCH gebildet werden [LfU, 1993; Engst et al, 1977]. Der Abbau
durch Bodenmikroorganismen erfolgt am leichtesten für γ-HCH. Die Abbauraten der
HCH-Isomere und auch die Abbaumechanismen sind unter aeroben und anaeroben
Bedingungen unterschiedlich. Unter anaeroben Bedingungen kann es je nach Isomer zu
einer Dechlorierung kommen [Haider, 1983]. Der mikrobiologische und physiko-
chemische Abbau findet in oberflächennahen Schichten statt. So können in
mikrobiologisch aktiven Böden 50 - 70 % der Substanz innerhalb eines Jahres abgebaut
werden [Koch, 1986]. α-Isomere werden wesentlich schneller aus dem Boden eliminiert
als β-Isomere. β-HCH zeigt von allen Isomeren die größte Persistenz im Boden.
Der Abbau von HCH-Isomeren aus dem Boden ist auf folgenden Wegen möglich:
• durch Einwirkung von Mikroorganismen (aerob und anaerob)
• durch chemische Reaktionen
• durch photochemische Reaktionen (obere Bodenschichten)
• durch Kodestillation
• durch Verdampfen
[Koch, 1986; Korte, 1983].
Durch den vergleichsweise hohen Dampfdruck sind die Stoffübergänge Wasser - Luft
und Boden – Luft zu beachten [Koch, 1986].
2.3 Aufnahme von organischen Umweltchemikalien in Pflanzen
Aufgrund der Bedeutung von Chemikalienrückständen in pflanzlichen Nahrungsmitteln,
die Aufnahme durch Pflanzenfresser und eventuelle Anreicherung ist die Aufnahme von
Chemikalien in Pflanzen von höchstem Interesse.
Page 9
2 HCH in der Umwelt
9
Die Aufnahmewege von Substanzen aus dem Boden in die Pflanze sind recht komplex.
Es gibt mehrere Aufnahme- und Verteilungswege von Chemikalien aus dem Boden in die
Pflanze:
• Aufnahme durch die Wurzel und Translokation in den Spross mit dem
Transpirationsstrom
• Aufnahme von aus dem Boden verflüchtigten Chemikalien über die Blätter
• Aufnahme von Substanzen aus Boden- und Staubpartikeln durch die Blätter
[Korte, 1992].
Die größte Bedeutung jedoch kommt bei Bodenkontaminationen der Aufnahme über die
Wurzel zu.
In welchem Maß Umweltchemikalien von der Pflanze aufgenommen werden, hängt unter
anderem von substanzspezifischen Parametern, wie Wasserlöslichkeit, Dampfdruck und
Kow (n-Oktanol/ Wasser-Verteilungskoeffizient) ab [Heinrich, 1997]. Die Aufnahme einer
Substanz über die Wurzel korreliert direkt mit ihrer pflanzenverfügbaren Form, d.h. mit
ihrer Konzentration im Bodenwasser [Korte, 1992; LfU, 1993]. Der Übergang der gelösten
Chemikalie in die Wurzel korreliert direkt mit dem n – Oktanol/ Wasser-Verteilungs-
koeffizienten, da unpolare Stoffe leichter aus der Bodenlösung an den Wurzeloberflächen
adsorbiert werden als polare und damit besser für eine Aufnahme in die Pflanze zur
Verfügung stehen. Die Translokation der aufgenommenen Stoffe innerhalb der Pflanze
verläuft für Substanzen mit mittlerer Polarität am leichtesten [Korte, 1992].
Weitere Einflussfaktoren für die Aufnahme einer Verbindung durch die Pflanzenwurzel
sind
• das Sorptionsvermögen des Bodens, das durch bodenspezifische Parameter wie
z. B. Gehalt an organischer Substanz und mineralischer Zusammensetzung
bestimmt wird
• die chemische Stabilität bzw. biologische Abbaubarkeit der Verbindung, die z. B.
durch pH-Wert und mikrobielle Aktivität des Bodens beeinflusst werden
• pflanzliche Charakteristika wie die Metabolisierung der Substanz in der Pflanze
• die Beschaffenheit des Wurzelsystems und der Blätter
• sowie die klimatischen Bedingungen wie Licht, Temperatur und Feuchtigkeit.
[Heinrich, 1997].
Zur quantitativen Beschreibung der relativen Anreicherung einer Substanz in der Pflanze
gegenüber dem Boden kann man den Transferfaktor (TF) verwenden. Dieser wird nach
folgender Formel berechnet:
TF = Schadstoffkonzentration in der Pflanze
Schadstoffkonzentration im Boden
Page 10
2 HCH in der Umwelt
10
Der TF wird auf die Trockenmasse bezogen. Der TF kann jedoch nur als grobe
Abschätzung dienen, da die Aufnahmeraten sowohl für den pflanzlichen Genotyp (Art,
Sorte) spezifisch sind, als auch von Boden- und Klimafaktoren (z. B. Bodenfeuchte,
Temperatur, Gehalt an organischer Substanz) abhängen. Der TF kann während einer
Vegetationsperiode einer Pflanze große Unterschiede aufweisen. Zur Beurteilung des
Transferfaktors müssen deswegen Angaben zu den Versuchsbedingungen und dem
physiologischen Alter der Pflanze mit betrachtet werden. Außerdem ist zu beachten, dass
in Gefäßversuchen die Aufnahmeraten überschätzt werden können, da hier ein größeres
Wurzel : Bodenverhältnis vorliegt als unter Freilandbedingungen [LfU, 1998].
Transferfaktoren kleiner 1 bedeuten, dass keine Bioakkumulation stattfindet [Heinrich,
1997].
Pflanzliche Metabolite von Umweltchemikalien sind vorwiegend Oxidationsprodukte.
Somit muss bei deren Auftreten mit einer gesteigerten Aktivität (Epoxide, Phenole) im
Boden und auch in Nahrungsmitteln gerechnet werden. Es kann aber auch zu einer
kovalenten Bindung oder einem nicht kovalentem Einschluss von Umweltchemikalien
oder ihrer Metabolite an bzw. in Makromoleküle (z. B. Lignin in Pflanzen oder
Huminstoffe im Boden) und somit zu einer temporären oder permanenten Fixierung/
Immobilisierung der Verbindung kommen [Heinrich, 1997].
2.4 Aufnahme und Metabolismus von HCH in Pflanzen
Die HCH-Isomere (α-, β- und γ-HCH) werden nach einer Spritzbehandlung von den
grünen Pflanzenteilen praktisch nicht resorbiert. In Abhängigkeit von der Kultur jedoch
kann eine Resorption über die Wurzel aus HCH-kontaminierten Böden eintreten. Vor
allem gilt dies für Wurzelgemüse, die im Allgemeinen über einen höheren Gehalt an
lipophilen Stoffen als andere Pflanzen verfügen [Eichler; 1983].
Der Boden stellt mit einer Halbwertszeit von 8-10 Jahren für β-HCH und über einem Jahr
für α- und γ-HCH die wesentliche Expositionsquelle für Pflanzen dar [Marquardt &
Schäfer, 1994]. Auf stark mit HCH kontaminierten Böden werden von den angebauten
Pflanzen beträchtliche Mengen HCH aufgenommen, wobei zwischen den HCH-Gehalten
im Boden und den HCH-Gehalten der Pflanzen, in Abhängigkeit von der Pflanzenart,
signifikante Korrelationen bestehen. Die HCH - Gehalte in den einzelnen Pflanzenteilen
steigen in folgender Reihenfolge: Korn < Frucht < Wurzel < Spross [Scheffer &
Schachtschabel, 2002].
An Tabakpflanzen wurde gezeigt, dass die Resorption für α- und γ-HCH größer ist als für
β- und δ-HCH [Kawahara & Nakamura, 1971]. Gao et al. (2005) untersuchten
verschiedene Gemüsearten u. a. auf HCH. Dabei wurden in Möhren, Rettich, Kopfsalat,
Page 11
2 HCH in der Umwelt
11
Kohl, Sellerie, Porree, Kürbis, Spinat und auch in Tomaten α-, β-, δ- und γ-HCH
nachgewiesen. Auch Tao et al. (2005) fanden in verschiedenen Gemüseproben HCH
(z.B. in Kohl, Spinat, Blumenkohl, Möhren). Gonzalez et al. (2003) stellten fest, dass die
HCH – Gehalte in Tomatenpflanzen im Laufe der Vegetationsperiode in Blättern,
Stängeln und Wurzel abnehmen, in Fruchtfleisch jedoch ansteigen. Dies könnte ein
Hinweis darauf sein, dass HCH die Fähigkeit zur Verlagerung innerhalb der
Pflanzengewebe und zur Akkumulation in Früchten besitzt.
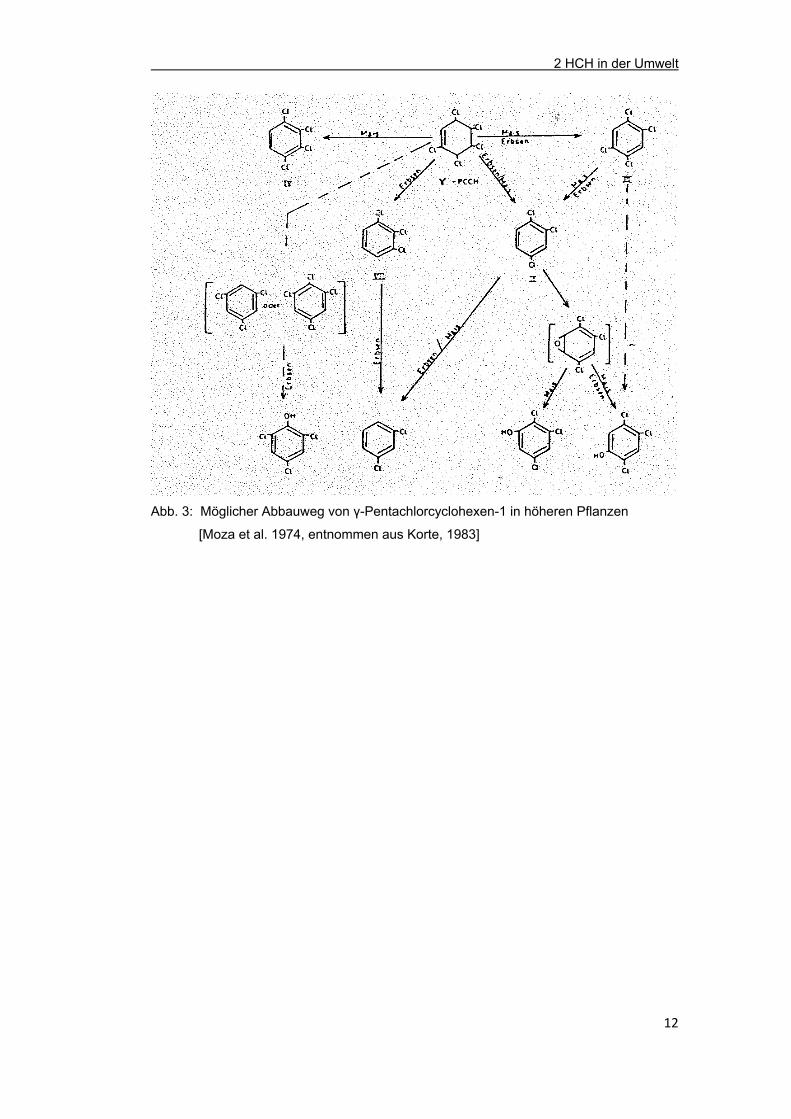
Als primäres Abbauprodukt von γ-HCH in Pflanzen kann 1,3,4,5,6-Pentachlorcyclohexen-
1 auftreten, das unter Chlorwasserstoffabspaltung in isomere Tri- und Tetrachlorbenzole
umgewandelt werden kann. Außerdem wurden neben Spuren verschiedener chlorierter
Benzole auch polare Metabolite wie Penta-, Tetra- und Trichlorphenole (frei oder in
konjugierter Form) in der Literatur erwähnt (Abb. 3) [Korte, 1983, LfU, 1993; DFG, 1982].
Bei den Reaktionsmechanismen handelt es sich um Dehydrochlorierung, Dehydrierung
und Dechlorierung. Außerdem kann im pflanzlichen Stoffwechsel auch eine
Hydroxylierung von Intermediärprodukten vorkommen. Dabei können die zunächst
gebildeten chlorierten Phenole in Glykoside umgewandelt werden.
Die Angaben zum Metabolismus in Pflanzen sind in der Literatur nicht einheitlich.
Allgemein kann jedoch gesagt werden, dass aufgrund meist relativ geringer
Konzentrationen in Pflanzen, der nicht sehr ausgeprägten Neigung zur Metabolisierung
im pflanzlichen Stoffwechsel und der Verdampfbarkeit, in Abhängigkeit von der Kultur
keine oder nur in untergeordnete Mengen Metabolite gefunden werden. Eine
Isomerisierung in höheren Pflanzen konnte nicht nachgewiesen werden [Eichler, 1983].
Dies bestätigen auch Untersuchungen zur Lindanaufnahme in Weidelgras. H. Li et al.
(2002) zeigten, dass es zu einem langsamen Anstieg der Lindankonzentration in den
Versuchspflanzen kommt bis eine Plateauphase erreicht wird. Dies deutet darauf hin,
dass der Metabolismus und die Bildung von gebundenen Rückständen kaum eine Rolle
spielen.
Page 12
2 HCH in der Umwelt
12
Abb. 3: Möglicher Abbauweg von γ-Pentachlorcyclohexen-1 in höheren Pflanzen
[Moza et al. 1974, entnommen aus Korte, 1983]
Page 13
3 HCH im Organismus
13
3 HCH im Organismus 3.1 Aufnahme, Verteilung, Metabolismus und Ausscheidung (Elimination) von HCH durch den Menschen
HCH-kontaminierte Lebensmittel stellen den Hauptexpositionsweg für den Menschen dar.
Zu über 90 % nimmt der Mensch Hexachlorcyclohexan mit der Nahrung auf. Die
Resorption über den Magen-Darm-Trakt erfolgt rasch und nahezu vollständig. Die
inhalative Resorptionsquote liegt bei ungefähr 50 % [Daunderer, 1991]. Für die dermale
Resorption liegen die Angaben zischen 10 % [WHO, 1991] und 40 % [LfU, 1993]. Hohe
Resorptionsquoten werden durch Lösung von HCH in fetthaltigen Trägermaterialien
erzielt [Marquardt & Schäfer, 1994].
Die Verteilung nach einmaligen Applikation ist offenbar nach 24 Stunden abgeschlossen
und erfolgt vor allem ins Fettgewebe und in lipidreiche Organe [LfU, 1993; Marquardt &
Schäfer, 1994]. Im Vergleich zum Blut werden in der Leber etwa 10-fach höhere und im
Zentralen Nervensystem (ZNS) etwa 3- bis 4-fach höhere Gehalte gefunden [Marquardt
& Schäfer, 1994].
Bei kurzfristiger Exposition, z.B. nach Aufbringung therapeutischer Dosen von γ-HCH auf
der Haut, wird es mit einer Halbwertszeit von etwa 20 Stunden aus dem Blut eliminiert.
Dagegen wird bei einer chronischen Exposition eine Halbwertszeit von 8 – 10 Tagen
beobachtet. Die Konfiguration des HCH-Isomers bestimmt die Geschwindigkeit der
Ausscheidung. Im Unterschied zu den anderen Isomeren wird β-HCH am langsamsten
metabolisiert und überwiegend mit dem Stuhl ausgeschieden. Beim α- und γ-HCH
überwiegt hingegen die Ausscheidung von Metaboliten. Beim γ-HCH wird nur ein kleiner
Teil unverändert über den Darm ausgeschieden [Marquardt & Schäfer, 1994]. Der
überwiegende Teil des Lindans hingegen wird durch Cytochrom-P450-abhängige
Monooxygenasen und Glutathiontransferasen der Leber metabolisiert. Dabei spielen die
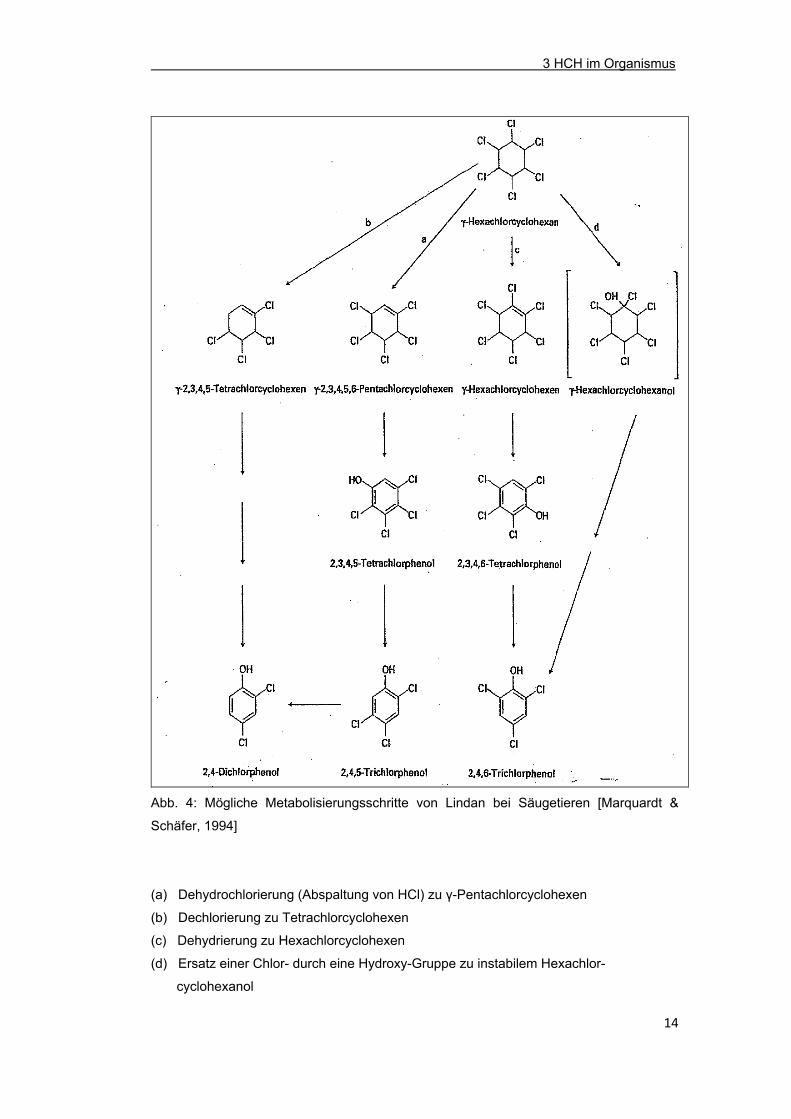
in Abb. 4 gezeigten Reaktionsschritte eine Rolle [Marquardt & Schäfer, 1994]. Die
Metaboliten werden hauptsächlich mit dem Harn eliminiert [Daunderer, 1991].
Page 14
3 HCH im Organismus
14
Abb. 4: Mögliche Metabolisierungsschritte von Lindan bei Säugetieren [Marquardt &
Schäfer, 1994]
(a) Dehydrochlorierung (Abspaltung von HCl) zu γ-Pentachlorcyclohexen
(b) Dechlorierung zu Tetrachlorcyclohexen
(c) Dehydrierung zu Hexachlorcyclohexen
(d) Ersatz einer Chlor- durch eine Hydroxy-Gruppe zu instabilem Hexachlor-
cyclohexanol
Page 15
3 HCH im Organismus
15
In weiteren Schritten werden die gebildeten Metaboliten zu Tetra- und Trichlorphenolen
abgebaut. Diese werden dann als Konjugate mit Glutathion, Glucuronsäure, Sulfat oder
Mercaptursäure ausgeschieden [Marquardt und Schäfer, 1994; Daunderer, 1991].
Erfahrungen am Menschen haben gezeigt, dass hohe Konzentrationen in der Luft (z.B.
am Arbeitsplatz) zu einer Akkumulation von HCH im Organismus führen [Baumann et al.
1983]. Aufgrund der langsamen Eliminierung von β-HCH ist das Anreicherungspotential
für dieses Isomer am größten. Es hat eine 10 bis 30fach größere Neigung im Fettgewebe
zu akkumulieren als γ-HCH. Daher tritt das β-Isomer in menschlichen Fett- und
Milchproben in den höchsten Konzentrationen auf [Jensen, 1983]. Lindan überwindet die
Plazentaschranke [ATSDR, 2005].
3.2 Akute Toxizität
HCH-Isomere beeinflussen an Säugetieren Funktionen des Zentralen Nervensystems.
Die chemische Erregbarkeit des ZNS wird herabgesetzt [Portig & Vohland, 1983].
Im Tierversuch wurde gezeigt, dass γ-HCH akut toxischer als α-, β- und δ-HCH ist [LfU,
1993].
Die Symptomatik der akuten Lindan-Vergiftung entspricht der anderer chlorierter
cyclischer Kohlenwasserstoffe [Marquardt & Schäfer, 1994]. HCH-Isomere wirken
neurotoxisch an Säugetieren [LfU, 1993]. Jedes der vier Isomere α – δ besitzt seine
eigene Form dieser Neurotoxizität (Tab. 2), wobei die nicht insektizid wirksamen HCH-
Isomere tierexperimentell nicht so intensiv geprüft wurden wie das γ-HCH.
Tab. 2: HCH-Vergiftung der Ratte: Neurotoxische Zustandsbilder [Portig & Vohland,
1983]
Isomer Prominente Symptome
α
β
γ
δ
Persistierender, generalisierter grober Tremor
Störung der motorischen Koordination (Ataxie) und Verlust der
groben Muskelkraft (Adynamie)
Repetitive klonisch-tonische Krämpfe
Verlust der Stell- und Haltereflexe („Narkose“)
Alle diese Vergiftungsbilder können, bei genügend hoher Dosis, tödlich enden. Der Tod
tritt innerhalb von Minuten (γ), Stunden (δ) oder Tagen (α und β) ein [Portig et al. 1983].
Die Leitsymptome einer Lindan-Intoxikation sind Reaktionen des zentralen Nerven-
systems [Herbst, 1983]. Nerven werden bei geringerer Konzentration Lindan
Page 16
3 HCH im Organismus
16
übererregbar, bei hoher Konzentration gelähmt. Die motorischen Bahnen sind zuerst
betroffen, bei hohen Konzentrationen auch spinale Bahnen [Daunderer, 1991].
Bereits nach 15 – 17 mg Lindan/kg Körpergewicht (KG) (orale Aufnahme) treten schwere
Intoxikationen auf, so dass 10 – 20 mg/kg KG lebensgefährlich wirken können.
Zentralnervöse Symptome, wie Übelkeit, Unruhe, Kopfschmerz, Erbrechen,
Muskelzuckungen, Gleichgewichtsverlust und tonisch-klonische Krämpfe, treten Minuten
bis Stunden nach der Exposition auf. Todesursache ist zentrales Atemversagen oder
Kreislaufkollaps [Herbst, 1983; WHO, 1991]. Die Übererregbarkeit des ZNS ist
wahrscheinlich das Resultat der antagonistischen Wirkung von Organochlorverbindungen
an den Rezeptoren der γ-Aminobuttersäure (GABA) im Gehirn. Damit wird der durch
GABA vermittelte Chloridstrom in die Zelle unterbunden, so dass das innen negative
Ruhemembranpotential nicht aufrechterhalten werden kann. Außerdem wird die calcium-
und magnesiumabhängige ATPase an der Plasmamembran der Nervenzellen gehemmt.
Die dadurch erhöhten Calciumkonzentrationen führen zu einer verstärkten
Neurotransmitterfreisetzung des präsynaptischen Neurons und tragen damit zu der
allgemeinen Übererregbarkeit des ZNS bei [Dekant & Vamvakas, 1994].
Abb. 5: Wirkungsmechanismus der insektiziden Organochlorverbindungen [Dekant &
Vamvakas, 1994]
Lindan selbst wird als Ursache der Krampfauslösung angesehen, d. h. es ist keine
vorherige Metabolisierung nötig [LfU, 1993]. Bereits 30 min nach oraler Aufnahme einer
toxischen Dosis Lindan wurden beim Menschen Krämpfe beobachtet. Auch nach
dermaler Applikation von 10 mg γ-HCH/kg KG traten diese Symptome auf [DFG, 1982].
α-HCH stimuliert das ZNS geringer und löst bei akuter Vergiftung (Ratte) einen
langanhaltenden, generalisierten Tremor aus. β-HCH hingegen vermindert die ZNS-
Aktivität. Ratten verlieren die grobe Muskelkraft und die Fähigkeit zur koordinierten
Page 17
3 HCH im Organismus
17
Bewegung. Todesursache ist unter anderem die Unfähigkeit, Nahrung oder Trinkwasser
aufzunehmen. Die Induktion hepatischer Enzymsysteme setzt nach mehreren Stunden
bis wenigen Tagen ein. Es kommt durch Zellvergrößerung oder Zellvermehrung zu einer
Lebervergrößerung. α-HCH erhöht in der Leber die DNA-Synthese- und die Mitoserate
[Marquardt & Schäfer, 1994].
Alle vier Isomere senkten im Tierversuch die Wirksamkeit zahlreicher Krampfgifte. Dieser
antikonvulsive Effekt beruht zum einen auf einer induktiven Erhöhung der Aktivität
fremdstoffmetabolisierender Enzyme, zum anderen können die HCH-Isomere auch durch
direkte Einwirkungen auf das ZNS dessen chemische und elektrische Erregbarkeit
herabsetzen [LfU, 1993]. Die α-, β- und δ-Isomere zeigen den antikonvulsiven Effekt
bereits in Dosierungen, die 3 – 5fach unter den neurotoxisch wirksamen Mengen liegen.
Bei γ-HCH jedoch treten antikonvulsive und neurotoxische Wirkungen bei ungefähr
gleicher Konzentration ein [Herbst & Bodenstein, 1973].
Die akute orale Toxizität von α- und β – HCH bei Ratten und Mäusen (LD50 = 0,5 – 4,5
g/kg bzw. 8 – 16 g/kg KG) ist wesentlich geringer als die von γ – HCH (LD50 = 60 – 250
mg/kg KG). Die dermale LD50 von Lindan beträgt bei Ratten ungefähr 900 mg/kg KG
[Marquardt & Schäfer, 1994].
3.3 Chronische Toxizität
Bei chronischer Fütterung von Ratten erwies sich β-HCH als giftiger als die α- und γ-
Isomere [Marquardt & Schäfer, 1994]. β-HCH ist zwar geringer akut toxisch als γ-HCH,
durch seine besonderen physikalisch-chemischen und pharmakokinetischen
Eigenschaften jedoch kann die höhere Toxizität des β-HCH bei wiederholter Applikation
erklärt werden [LfU, 1993; Portig et al. 1983].
Nach niedriger Dosierung wurde bei Ratten, Mäusen und Hamstern ein erhöhtes
Lebergewicht beobachtet [Marquardt & Schäfer, 1994; LfU, 1993], da die HCH-Isomere
mikrosomale Enzyme der Leber induzieren [Hoffmann, 1983]. Nach hohen Dosen zeigten
sich eine fettige Degeneration und fokale Nekrose der Leberzellen, schließlich chronische
Nephritis und Lebenszeitverkürzung. Für α- und β-HCH wurde ein NOAEL von 0,1 mg/kg
und für γ-HCH von 0,2 – 2 mg/kg KG abgeleitet. In einer Inhalationsstudie an Ratten
wurde ein NOAEL von 0,6 mg Lindan/ m3 geschätzt [Marquardt & Schäfer, 1994].
Offenbar gehört HCH in die Gruppe körperfremder Substanzen, die im Knochenmark des
Menschen die Regeneration aller Blutzellen oder Zellreihen inhibieren können. Nach
intensivem Kontakt mit unterschiedlichen HCH-Isomeren wurden beim Menschen
Page 18
3 HCH im Organismus
18
mehrfach isolierte Thrombozytopenien oder aplastische Anämien, manchmal
Panmyelophtisen beschrieben. Die Erkrankungen führten oft zum Tode. Eine Beziehung
zur eingenommenen Dosis oder zur Expositionsdauer konnte nicht festgestellt werden
[Vohland et al. 1983].
Beim Menschen können ab einer Blutkonzentration von 0,02 mg/l klinische, insbesondere
zentralnervöse Symptome auftreten. Enzyminduktionen, wie schnellere Chininexkretion
und schnellere Antipyrin- und Phenylbutazor-Ausscheidung, sind nach höherer
beruflicher Exposition bekannt. Sehr vereinzelt treten allergische Reaktionen auf [Herbst,
1983].
3.4 Spezielle Toxikologie
Reproduktionstoxizität
Lindan sowie die anderen HCH-Isomere besitzen keine teratogene Wirkung [Marquardt &
Schäfer, 1994]. Die Substanz ist jedoch plazentagängig [ATSDR, 2005].
Nach oraler Zufuhr von 5 mg/kg KG zeigten sich weder fetotoxische noch
maternaltoxische Wirkungen. Diese Dosis übte in einer Dreigenerationenstudie an Ratten
keine reproduktionstoxische Wirkung aus. In der 3. Generation wurde eine
Enzyminduktion beobachtet. Eine Dosis von 0,5 mg β-HCH/ kg KG führte in einer
Zweigenerationenstudie an Ratten zu erhöhter Sterblichkeit und Infertilität. Eine Dosis
von 0,1 mg/kg zeigte keine Effekte [Marquardt & Schäfer, 1994].
Die dermale Applikation von technischem HCH an Ratten verursachte eine Abnahme der
Testosteronkonzentration im Serum, der Spermienzahl sowie der Spermienbeweglichkeit.
Außerdem nahm die Zahl der abnormen Spermien zu und es waren veränderte
Aktivitäten von Hodenenzymen feststellbar. Weibliche Ratten, die mit γ-HCH behandelt
wurden, zeigten signifikante Reduzierung der Ovulationsraten [Willett, 1998].
Herbst (1983) berichtet von Fetotoxizität bei der Maus (6 mg/kg, subcutan) und erhöhten
Totgeburten bei Hunden. Es gibt auch Hinweise auf Schäden während der
Kindesentwicklung nach der Geburt durch γ-HCH [Witte et al. 1988].
Mutagenität, Kanzerogenität Die HCH-Isomere sind nicht mutagen [Marquardt & Schäfer, 1994; Hoffmann, 1983].
Lindan verursacht an pflanzlichen und tierischen Zellen colchicinähnliche
Mitosehemmung. α-, β- und γ-HCH erzeugen Polyploidie in tierischen und pflanzlichen
Zellen. Außerdem wurden Chromosomenbrüche und –lücken in Geweben von
Versuchstieren und in menschlichen Lymphozytenkulturen beobachtet [DFG, 1982]. In
Page 19
3 HCH im Organismus
19
menschlichen Lymphozytenkulturen konnten ab 0,5 µg γ-HCH/ml Medium dosis-
abhängige Chromatidbrüche beobachtet werden [Herbst, 1983; DFG, 1982]. Jedoch stellt
Lindan für den Menschen kein genetisches Risiko dar [Herbst, 1983].
Im Langzeitversuch zeigte α-HCH bei Mäusen ab einer Dosis von 100 – 250 mg/kg
Futter, bei Ratten ab 1000 mg/kg Lebertumoren. Nach Marquardt und Schäfer (1994)
zeigten β- und γ- HCH unter diesen Bedingungen keine kanzerogene Wirkung. Schulte-
Herrmann (1983) beobachtete aber auch nach β- und γ-HCH-Behandlung in bestimmten
Mäusestämmen Tumore oder Tumorvorstufen.
Bei Mäusen scheint es einen „Schwellenwert“ im Bereich von 300 mg γ-HCH/kg Futter
(entspricht ca. 43 mg/kg KG) zu geben, bis zu dem in der Mäuseleber keine erhöhte
Hepatom- bzw. Carcinomrate bei den behandelten Tieren gefunden wurde. Vermehrtes
Lebertumorwachstum konnte nur nach Langzeitwirkung akut toxischer Dosierungen
festgestellt werden [Herbst, 1983].
α-HCH induzierte im Tumorpromotionsversuch mit initialer Gabe von Diethylnitrosamin
eine verstärkte DNA-Syntheserate in γ-GT-positiven Leberfoci. Es entwickelten sich
größere Lebertumoren als in den Vergleichsratten. Die Überlebenszeit war verkürzt. β-
HCH gilt ab einer Dosis von 3 mg/kg KG als Tumorpromotor [Marquardt & Schäfer,
1994]. Die Tumorinduktion durch γ-HCH erfolgt durch kovalente Bindung von
Chemikalien an die DNS [Herbst, 1983]. Aufgrund dieser Befunde wird HCH
(insbesondere α- und γ-HCH) als potentiell kanzerogener Stoff angesehen [DFG, 1982].
Die US – Umweltschutzbehörde EPA stufte α–HCH als wahrscheinlich krebserregend, β-
HCH als möglicherweise krebserregend und γ-HCH als nachweislich krebserregend ein
[EPA, 2000].
Immuntoxizität
Lindan kann Teilfunktionen der humoralen und zellulären Abwehr unterdrücken
[Marquardt & Schäfer, 1994]. In vitro Experimente mit γ-HCH (0,5 µg/ml Medium) zeigten
eine Hemmung der spontanen Transformation von Lymphozyten zu Lymphoblasten
sowie der Aufnahme von Antigen-Antikörper-Komplexen von der Zelloberfläche in das
Zellinnere. Bei Konzentrationen über 5 µg γ-HCH/ml Medium waren zytotoxische
Wirkungen (z.B. morphologische Veränderungen) erkennbar. Während die γ- und δ-
Isomere die Proliferation von Lymphozyten hemmten, konnte eine ähnliche Wirkung
selbst bei hohen α- und β-HCH-Konzentrationen nicht nachgewiesen werden. Bei 60
Beschäftigten aus der HCH-Produktion wurden nach 10jähriger Exposition im peripheren
Blut etwa 20 % weniger Lymphozyten gezählt als bei einem Kollektiv nicht-exponierter,
gleichaltriger Personen [Vohland & Koransky, 1983].
Page 20
4 Material und Methoden
20
4 Material und Methoden
4.1 Versuchsdurchführung
Da Nutzpflanzen in der Lage sind, Schadstoffe aus dem Boden aufzunehmen und
anzureichern [z.B.: Verma, 1991, Heinrich, 1997, Gonzalez et al. 2003] und der Haupt-
aufnahmeweg von Lindan durch den Menschen über kontaminierte Lebensmittel erfolgt
[Marquardt & Schäfer, 1994], resultiert über die Nahrungskette ein Gefährdungspotential
für den Menschen.
In der vorliegenden Arbeit werden Nutzpflanzen mit Lindan exponiert, um das
Transferverhalten in die Pflanze und gegebenenfalls in die Früchte zu untersuchen. Bei
den Versuchspflanzen handelt es sich um Tomatenpflanzen (Lycopersicon esculentum),
die im Gewächshaus unter kontrollierten Bedingungen kultiviert wurden. Die Experimente
wurden als Gefäßversuche (4-fach Bestimmung) durchgeführt. Um dabei den Einfluss
von Bodenfaktoren (z.B. organische Substanz) weitestgehend auszuschließen, wurden
die Pflanzen in einer Sand-Hydrokultur angebaut, wobei das Nährlösungsvolumen (2,5l)
durch tägliches Gießen konstant gehalten wurde. Die Testsubstanz Lindan wurde mit der
Nährlösung zugegeben. Jede Pflanze wurde über den Zeitraum einer Vegetationsperiode
mit 3 mg Lindan versetzt. Dies entspricht einer Bodenkonzentration von 1 mg/kg.
Nach der Probenahme der Pflanzen wurden diese in ihre ober- und unterirdischen
Pflanzenteile getrennt. Dabei wurde der oberirdische Pflanzenteil in 0 – 50 cm und > 50
cm unterteilt. Die Früchte der Tomatenpflanzen wurden bereits während des Wachstums
bei erreichtem Reifezustand geerntet. Die Wurzeln wurden gründlich mechanisch
gereinigt, um anhaftenden Bodenbestandteilen zu entfernen und anschließend kurz mit
bidestilliertem Wasser gewaschen, um die Nährlösung zu entfernen. Nach der
Separierung der Pflanzen erfolgte die Bestimmung der Frischmasse. Das
Pflanzenmaterial wurde bis zur Analyse bei -20°C gelagert.
Weiterhin wurden Nährlösungen und Böden (Sand) beprobt.
Zunächst wurde das Volumen der Nährlösungen bestimmt und bis zur weiteren
Verwendung im Kühlschrank gelagert.
Nach Homogenisierung des Bodenmaterials (3 kg je Pflanze) wurde eine repräsentative
Teilprobe entnommen und luftgetrocknet. Frisch- und Trockengewicht der Böden wurden
bestimmt.
Page 21
4 Material und Methoden
21
4.2 Analytik
4.2.1 Probenvorbereitung
Nährlösung
Die Probenvorbereitung erfolgte mittels Festphasenmikroextraktion (SPME, Abb. 6). Das
Prinzip der SPME beruht auf der Anreicherung von Analyten auf einer
polymerbeschichteten Quarzfaser durch direktes Eintauchen der Faser in die Probe. Die
maximale Extraktion der Analyten wird nach Erreichen des Verteilungsgleichgewichtes
der Analyten zwischen wässriger Phase und SPME-Faserbelegung erzielt. Anschließend
werden die angereicherten Moleküle thermisch im Injektor eines Gaschromatographen
(GC) desorbiert und mittels GC analysiert. Die SPME ist eine komplett lösemittelfreie
Extraktionstechnik und verbindet Analytanreicherung und Probenaufgabe in einem
Probenvorbereitungsschritt. Sie ist automatisierbar und stellt somit eine einfache und
zeitsparende Methode dar [Pawliszyn, 1997].
Abb. 6: SPME – Faser
Mit folgenden SPME-Bedingungen wurde nach der Optimierung gearbeitet:
• Faserbeschichtung: 85 µm Polyacrylat (Abb. 7)
• Anreicherungszeit: 60 min
• Anreicherungsart: Immersion
Page 22
4 Material und Methoden
22
• Rührgeschwindigkeit: 750 U/min
• Probenvolumen: 9 ml
• Salzzugabe: 2,25 g NaCl/ 9 ml Probe
• Desorptionszeit: 3 min
• Desorptionstemperatur: 280 °C.
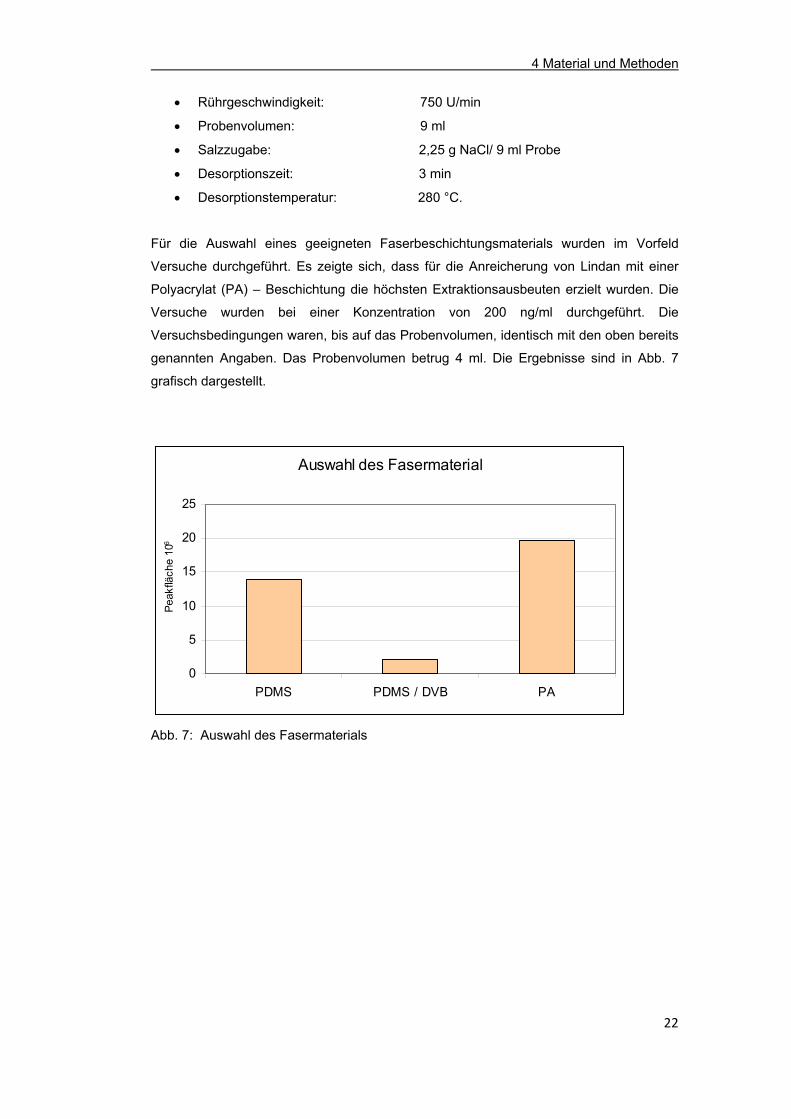
Für die Auswahl eines geeigneten Faserbeschichtungsmaterials wurden im Vorfeld
Versuche durchgeführt. Es zeigte sich, dass für die Anreicherung von Lindan mit einer
Polyacrylat (PA) – Beschichtung die höchsten Extraktionsausbeuten erzielt wurden. Die
Versuche wurden bei einer Konzentration von 200 ng/ml durchgeführt. Die
Versuchsbedingungen waren, bis auf das Probenvolumen, identisch mit den oben bereits
genannten Angaben. Das Probenvolumen betrug 4 ml. Die Ergebnisse sind in Abb. 7
grafisch dargestellt.
Auswahl des Fasermaterial
0
5
10
15
20
25
PDMS PDMS / DVB PA
Peak
fläch
e 10
6
Abb. 7: Auswahl des Fasermaterials
Page 23
4 Material und Methoden
23
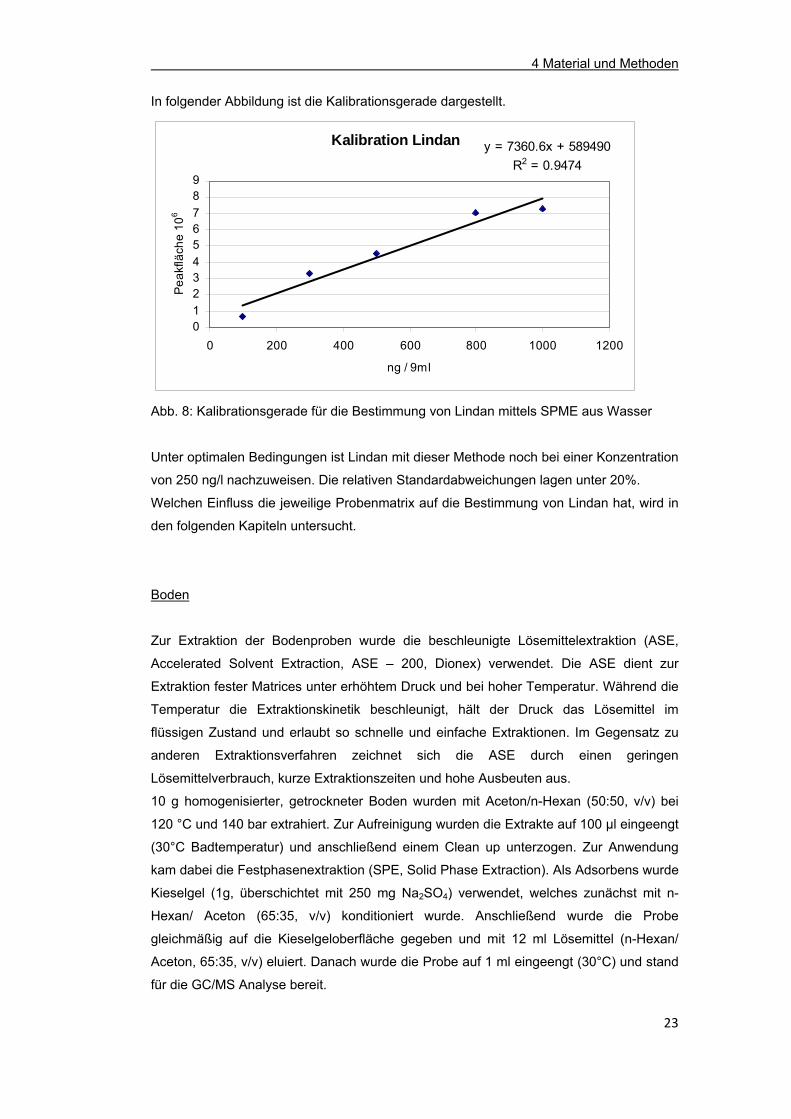
In folgender Abbildung ist die Kalibrationsgerade dargestellt.
Kalibration Lindan y = 7360.6x + 589490R2 = 0.9474
0123456789
0 200 400 600 800 1000 1200
ng / 9ml
Pea
kflä
che
106
Abb. 8: Kalibrationsgerade für die Bestimmung von Lindan mittels SPME aus Wasser
Unter optimalen Bedingungen ist Lindan mit dieser Methode noch bei einer Konzentration
von 250 ng/l nachzuweisen. Die relativen Standardabweichungen lagen unter 20%.
Welchen Einfluss die jeweilige Probenmatrix auf die Bestimmung von Lindan hat, wird in
den folgenden Kapiteln untersucht.
Boden
Zur Extraktion der Bodenproben wurde die beschleunigte Lösemittelextraktion (ASE,
Accelerated Solvent Extraction, ASE – 200, Dionex) verwendet. Die ASE dient zur
Extraktion fester Matrices unter erhöhtem Druck und bei hoher Temperatur. Während die
Temperatur die Extraktionskinetik beschleunigt, hält der Druck das Lösemittel im
flüssigen Zustand und erlaubt so schnelle und einfache Extraktionen. Im Gegensatz zu
anderen Extraktionsverfahren zeichnet sich die ASE durch einen geringen
Lösemittelverbrauch, kurze Extraktionszeiten und hohe Ausbeuten aus.
10 g homogenisierter, getrockneter Boden wurden mit Aceton/n-Hexan (50:50, v/v) bei
120 °C und 140 bar extrahiert. Zur Aufreinigung wurden die Extrakte auf 100 µl eingeengt
(30°C Badtemperatur) und anschließend einem Clean up unterzogen. Zur Anwendung
kam dabei die Festphasenextraktion (SPE, Solid Phase Extraction). Als Adsorbens wurde
Kieselgel (1g, überschichtet mit 250 mg Na2SO4) verwendet, welches zunächst mit n-
Hexan/ Aceton (65:35, v/v) konditioniert wurde. Anschließend wurde die Probe
gleichmäßig auf die Kieselgeloberfläche gegeben und mit 12 ml Lösemittel (n-Hexan/
Aceton, 65:35, v/v) eluiert. Danach wurde die Probe auf 1 ml eingeengt (30°C) und stand
für die GC/MS Analyse bereit.
Page 24
4 Material und Methoden
24
ASE – Methode:
Lösemittel: Aceton / n-Hexan (50:50, v/v)
Druck: 140 bar
Temperatur: 120 °C
Pflanzen
Das Pflanzenmaterial, welches bis zur Analyse bei -20°C lagerte, wurde zunächst
aufgetaut, luftgetrocknet, die Trockenmasse bestimmt und anschließend gemahlen. Eine
repräsentative Teilprobe (ca. 5g) wurde entnommen und mittels Soxhlet-Extraktion 4
Stunden lang mit Cyclohexan extrahiert. Im Anschluss wurden die Extrakte filtriert und
auf 1 ml eingeengt. Für das Clean up wurde eine SPE- Kartusche mit 1 g Silicagel und
250 mg Natriumsulfat verwendet. Die Analyten wurden mit 10 ml n-Hexan/ Aceton (65/35,
v/v) eluiert. Die Extrakte wurden auf 1 ml eingeengt und mittels GC-MS analysiert.
Zusätzlich wurden während der Vegetationsperiode in vivo – Messungen mittels SPME in
den lebenden Pflanzen durchgeführt. Hierfür wurden die SPME-Fasern in den Stamm der
Pflanze eingebracht und zum gleichen Zeitpunkt Lindan zur Nährlösung gegeben. Die
SPME- Fasern wurden eine Woche lang in der Pflanze belassen und anschließend
mittels GC-MS analysiert.
Früchte
Die Früchte, welche wie die Pflanzenproben bei -20 °C lagerten, wurden aufgetaut,
püriert und in einem Verhältnis von 1:1 mit destilliertem Wasser verdünnt, um eine
optimale Durchmischung der Probe während der Anreicherung zu erzielen. Die
Anreicherung des Lindans aus der Fruchtmatrix erfolgte mittels SPME (Abb. 9). Da die
Probenmatrix die Extraktionsausbeuten der Zielsubstanz erheblich beeinflussen kann,
wurden in Vorversuchen mit dotierten Früchten die Extraktionsparameter variiert. Dabei
spielten vor allem die Extraktionszeit und die Verdünnung eine Rolle. Die optimierten, an
die Fruchtmatrix angepassten Parameter sind im Folgenden dargestellt:
• Verhältnis Probe:Wasser: 1:1
• Probenvolumen: 9 ml
• Anreicherungszeit: 60 min
• Faserbeschichtung: Polyacrylat
• Anreicherungsart: Immersion
• Rührgeschwindigkeit: 1000 U/min
Page 25
4 Material und Methoden
25
• Salzgehalt: 2,25 g
• Desorptionszeit: 3 min
• Desorptionstemperatur: 280 °C
Abb. 9: Anreicherung des Lindans aus der Fruchtmatrix mittels SPME
Trotz der optimierten Parameter wird die Extraktionsausbeute durch die Probenmatrix
beeinflusst. Die Wiederfindung für Lindan in der Fruchtmatrix lag bei 19% gegenüber
wässrigen Proben (Abb. 10).
Abb.10: Matrixabhängigkeit
Page 26
4 Material und Methoden
26
4.2.2 Analytik
Für die Analysen der Nährlösungen wurde die Gaschromatographie mit MS-Detektion
(HP 6890 GC-System und ein 5973 MSD, Agilent Technologies, Autosampler MPS2 (Fa.
Gerstel)) eingesetzt. Die Analysen der Boden- und Pflanzenextrakte wurden mit dem
GC-System 6890N und 5973N MSD (beide von Agilent Technologies) durchgeführt. Die
Analysenbedingungen werden aus Tab. 3 ersichtlich.
Tab. 3: Trennbedingungen bei den GC/MS Analysen
Trennsäule HP 5-MS (ID 0,25 mm, Länge 30m,
Film 0,25 µm)
Injektor Splitless, 280 °C
Trägergas Helium
Temperaturprogramm
40 °C (1 min)
15 K/min bis 250 °C
250 °C (2 min)
Analysenzeit 17 min
Detektortemperatur = Ionenquellentemperatur
Transferline -Temperatur
180 °C
280 °C
Scan-Modus Single Ion Monitoring (SIM)
Page 27
5 Ergebnisse
27
5 Ergebnisse
5.1 Rückstandsanalytik in der Nährlösung und im Boden
In der folgenden Tabelle sind die am Versuchsbeginn zugegebenen Konzentrationen
Lindan sowie die nach einer Vegetationsperiode ermittelten Rückstände in der
Nährlösung und im Boden dargestellt.
Tab. 4: Lindan-Konzentrationen zu Versuchsbeginn sowie Rückstände in Boden und
Nährlösung nach einer Vegetationsperiode
Tomate 1 Tomate 2 Tomate 3 Tomate 4 Kontroll-
pflanze
Zugegebene
Lindan-Konz.
[µg/kg]
1000 1000 1000 1000
n.n.
Nährlsg. [µg/l] 2,85 5,12 1,09 2,17 n.n.
Boden [µg/kg] 11,0 7,0 17,0 12,0 n.n. n.n. nicht nachweisbar
Bei den 4 Versuchsansätzen mit Schadstoffzugabe (Tomate 1 bis 4), sind nach der Ernte
noch Lindanrückstände im Boden und in der Nährlösung nachweisbar. Es wurde parallel
eine Kontrollpflanze mitgeführt, bei der keine Schadstoffzugabe erfolgte. Hier waren
weder in der Nährlösung noch im Boden Lindanrückstände zu finden.
5.2 Lindankonzentrationen in den Versuchspflanzen
Die Pflanzen wurden getrennt nach Wurzel und oberirdischem Pflanzenteil analysiert. Bei
den Tomaten 1 – 3 wurde außerdem der oberirdische Pflanzenteil in den Bereich 0-50
cm und >50 cm unterteilt. Bei Tomate 4 wurde die oberirdische Biomasse nicht getrennt
untersucht.
In Tab. 5 sind die Lindankonzentrationen der Versuchspflanzen dargestellt.
Page 28
5 Ergebnisse
28
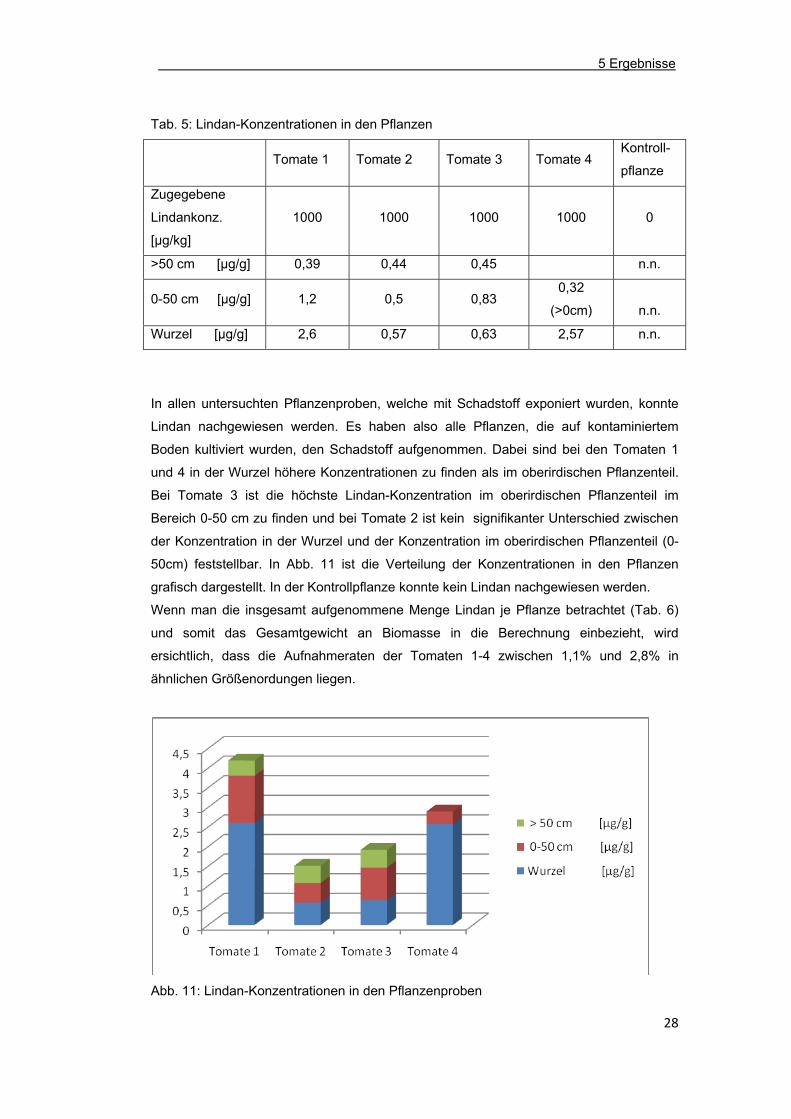
Tab. 5: Lindan-Konzentrationen in den Pflanzen
Tomate 1 Tomate 2 Tomate 3 Tomate 4
Kontroll-
pflanze
Zugegebene
Lindankonz.
[µg/kg]
1000 1000 1000 1000 0
>50 cm [µg/g] 0,39 0,44 0,45 n.n.
0-50 cm [µg/g] 1,2 0,5 0,83 0,32
(>0cm)
n.n.
Wurzel [µg/g] 2,6 0,57 0,63 2,57 n.n.
In allen untersuchten Pflanzenproben, welche mit Schadstoff exponiert wurden, konnte
Lindan nachgewiesen werden. Es haben also alle Pflanzen, die auf kontaminiertem
Boden kultiviert wurden, den Schadstoff aufgenommen. Dabei sind bei den Tomaten 1
und 4 in der Wurzel höhere Konzentrationen zu finden als im oberirdischen Pflanzenteil.
Bei Tomate 3 ist die höchste Lindan-Konzentration im oberirdischen Pflanzenteil im
Bereich 0-50 cm zu finden und bei Tomate 2 ist kein signifikanter Unterschied zwischen
der Konzentration in der Wurzel und der Konzentration im oberirdischen Pflanzenteil (0-
50cm) feststellbar. In Abb. 11 ist die Verteilung der Konzentrationen in den Pflanzen
grafisch dargestellt. In der Kontrollpflanze konnte kein Lindan nachgewiesen werden.
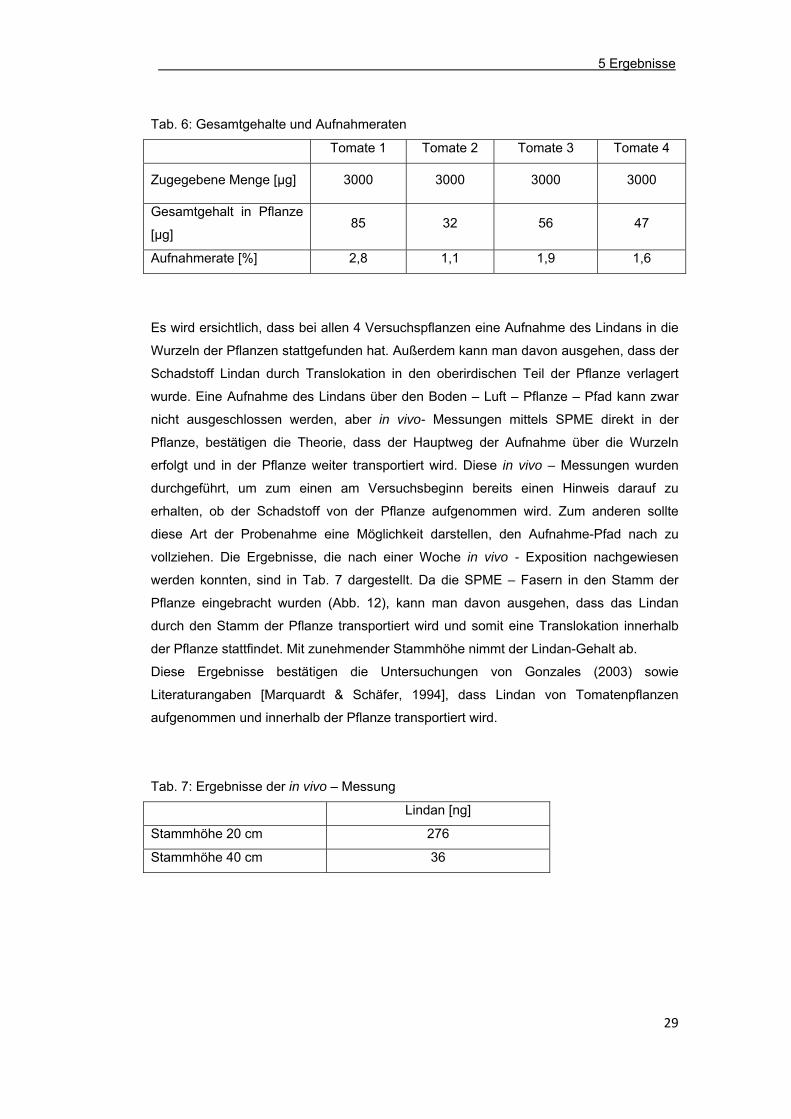
Wenn man die insgesamt aufgenommene Menge Lindan je Pflanze betrachtet (Tab. 6)
und somit das Gesamtgewicht an Biomasse in die Berechnung einbezieht, wird
ersichtlich, dass die Aufnahmeraten der Tomaten 1-4 zwischen 1,1% und 2,8% in
ähnlichen Größenordungen liegen.
Abb. 11: Lindan-Konzentrationen in den Pflanzenproben
Page 29
5 Ergebnisse
29
Tab. 6: Gesamtgehalte und Aufnahmeraten
Tomate 1 Tomate 2 Tomate 3 Tomate 4
Zugegebene Menge [µg] 3000 3000 3000 3000
Gesamtgehalt in Pflanze
[µg] 85 32 56 47
Aufnahmerate [%] 2,8 1,1 1,9 1,6
Es wird ersichtlich, dass bei allen 4 Versuchspflanzen eine Aufnahme des Lindans in die
Wurzeln der Pflanzen stattgefunden hat. Außerdem kann man davon ausgehen, dass der
Schadstoff Lindan durch Translokation in den oberirdischen Teil der Pflanze verlagert
wurde. Eine Aufnahme des Lindans über den Boden – Luft – Pflanze – Pfad kann zwar
nicht ausgeschlossen werden, aber in vivo- Messungen mittels SPME direkt in der
Pflanze, bestätigen die Theorie, dass der Hauptweg der Aufnahme über die Wurzeln
erfolgt und in der Pflanze weiter transportiert wird. Diese in vivo – Messungen wurden
durchgeführt, um zum einen am Versuchsbeginn bereits einen Hinweis darauf zu
erhalten, ob der Schadstoff von der Pflanze aufgenommen wird. Zum anderen sollte
diese Art der Probenahme eine Möglichkeit darstellen, den Aufnahme-Pfad nach zu
vollziehen. Die Ergebnisse, die nach einer Woche in vivo - Exposition nachgewiesen
werden konnten, sind in Tab. 7 dargestellt. Da die SPME – Fasern in den Stamm der
Pflanze eingebracht wurden (Abb. 12), kann man davon ausgehen, dass das Lindan
durch den Stamm der Pflanze transportiert wird und somit eine Translokation innerhalb
der Pflanze stattfindet. Mit zunehmender Stammhöhe nimmt der Lindan-Gehalt ab.
Diese Ergebnisse bestätigen die Untersuchungen von Gonzales (2003) sowie
Literaturangaben [Marquardt & Schäfer, 1994], dass Lindan von Tomatenpflanzen
aufgenommen und innerhalb der Pflanze transportiert wird.
Tab. 7: Ergebnisse der in vivo – Messung
Lindan [ng]
Stammhöhe 20 cm 276
Stammhöhe 40 cm 36
Page 30
5 Ergebnisse
30
Abb. 12: SPME – Faser im Stamm einer Tomatenpflanze zur in vivo – Messung
Für eine weitere Beurteilung der Anreicherung von Lindan in der Pflanze wird der
Transferfaktor (TF) herangezogen. Dieser wird, wie auf S. 9 bereits näher erläutert, nach
folgender Formel berechnet:
TF = Lindankonzentration in der Wurzel
Lindankonzentration im Boden
Der Transferfaktor wurde für die Wurzel berechnet und wird in Tab. 8 angegeben und mit
Daten aus der Literatur verglichen.
Tab. 8: Transferfaktoren Boden-Pflanze von γ-HCH
Pflanzenteil TF (berechnet) TF (Literaturangaben)
Wurzel (n=4) 6,4 12.3 – 3.2 (Gonzalez et al. 2003)
Ein TF >1 zeigt an, dass eine Bioakkumulation stattfindet (siehe S. 9). Mit einem
berechneten Wert von 6,4 zeigt die Wurzel ein starkes Anreicherungsvermögen.
Vergleicht man den berechneten TFWurzel mit Angaben aus der Literatur, so zeigt sich eine
gute Übereinstimmung. Die Spanne der TF ist auf die unterschiedlichen
Probenahmezeitpunkte innerhalb der Vegetationsperiode zurück zuführen. Mit
zunehmender Wachstumsdauer nimmt der TF ab. Die Transferfaktoren aus der Literatur
sind außerdem nicht direkt mit den in eigenen Experimenten ermittelten TF vergleichbar,
da andere Versuchsbedingungen vorlagen und der Transferfaktor neben der
Page 31
5 Ergebnisse
31
Pflanzenspezies auch von Faktoren wie Temperatur und Gehalt an organischer Substanz
im Boden abhängt.
5.3 Lindankonzentrationen in Früchten
Alle 4 Versuchspflanzen, die mit Lindan versetzt wurden, bildeten Früchte aus. Diese
wurden bei erreichtem Reifezustand geerntet und analysiert. Insgesamt wurden 38
Früchte, die von unterschiedlichen Stammhöhen und zu unterschiedlichen Zeitpunkten
geerntet wurden, untersucht. In 35 von 38 Früchten konnte kein Lindan nachgewiesen
werden. In 3 Fruchtproben wurde Lindan gefunden. Die Konzentrationen lagen, nach
Berücksichtigung des Matrixeinflusses, im Bereich von 83 – 318 ng/g, unabhängig von
Wachstumshöhe und Erntezeitpunkt.
Aufgrund ihrer Persistenz, ihres Vorkommens im Boden sowie der Möglichkeit zur
Anreicherung in der Nahrungskette werden Pestizidrückstände in Lebensmitteln durch
Grenzwerte in der Rückstands-Höchstmengen-Verordnung (RHmV) gesetzlich geregelt.
Da der Grenzwert für Lindan in Fruchtgemüse laut RHmV bei 1 mg/kg liegt, sind die
Konzentrationen der Fruchtproben aus der Versuchsreihe als unbedenklich einzustufen.
Im Gegensatz zu den Untersuchungen von Gonzalez (2003) konnte keine Akkumulation
von Lindan in den Früchten der Tomatenpflanzen festgestellt werden.
Page 32
6 Zusammenfassung
32
6 Zusammenfassung
Hexachlorcyclohexan (HCH) umfasst eine Gruppe von 8 Stereoisomeren, die nicht als
natürliche Substanz in der Umwelt vorkommen, sondern durch Photochlorierung von
Benzol hergestellt werden. Technisches HCH besteht aus einem Isomerengemisch, dass
aus 65-70% α-HCH, 10% β-HCH, 15% γ-HCH, 7% δ-HCH sowie weiteren HCH-Isomeren
in geringerer Konzentration besteht. Durch die Anwendung von Technischem HCH als
Schädlingsbekämpfungsmittel gelangten alle Isomere in die Umwelt. Da aber nur γ-HCH
eine insektizide Wirkung besitzt, wurde später Lindan, das zu >99% aus γ-HCH besteht,
eingesetzt.
Aufgrund der Persistenz, der Mobilität in der Umwelt, der Fähigkeit zur Bioakkumulation
sowie der potentiellen Toxizität und Kanzerogenität ist heute der Einsatz von HCH bzw.
Lindan bis auf wenige Ausnahmen europaweit verboten. Trotzdem sind die HCH-
Isomere, aufgrund ihrer Persistenz, immer noch in allen Medien (z. B. Boden, Sedimente,
Pflanzen, Muttermilch) detektierbar. Durch ihre lipophilen Eigenschaften neigen die HCH-
Isomere zur Akkumulation in der Nahrungskette. Die Hauptexpositionsquelle für den
Menschen stellen Nahrungsmittel dar.
Da bekannt ist, dass Nutzpflanzen organische Schadstoffe anreichern können, wurden in
der vorliegenden Arbeit Tomatenpflanzen in einem Modellsystem mit Sandboden auf
Nährlösung mit Lindan exponiert und der Transfer und die Verteilung innerhalb der
Pflanzen untersucht. Dabei wurde festgestellt, dass eine Lindanaufnahme sowie eine
Translokation innerhalb der Pflanzen statt findet. Die Aufnahmeraten lagen zwischen 1,1
und 2,8 % der Ausgangskonzentration.
Es wurde außerdem der Transferfaktor für die Wurzel der Tomatenpflanzen berechnet.
Mit einem TF von 6,4 zeigte die Wurzel ein Anreicherungsverhalten, da ein TF >1 auf
eine Bioakkumulation schließen lässt.
Eine Anreicherung in den Früchten konnte jedoch nicht festgestellt werden. Es wurde nur
in 3 von 38 Früchten Lindan nachgewiesen. Die höchste gemessene Konzentration lag
bei 318 ng/g und somit unter dem Grenzwert der RHmV von 1 mg/kg. Somit ist unter den
hier vorliegenden Versuchsbedingungen für den Verbraucher kein Risiko beim Verzehr
der Früchte zu erwarten.
Um die Ergebnisse zu bestätigen, sind weitere Tests, auch mit veränderten
Versuchsbedingungen, nötig. Vor allem sollte geprüft werden, wie sich das Aufnahme-
verhalten ändert, wenn mit höheren Ausgangskonzentrationen gearbeitet wird, da es in
der Literatur [LfU, 1998] Hinweise darauf gibt, dass die Konzentrationen im
Pflanzengewebe im positiven Zusammenhang mit den Bodenkonzentrationen stehen.
Page 33
Literaturverzeichnis
33
Literaturverzeichnis Agency for Toxic Substances and Disease Registry (2005): Public Health statement Hexachlorocyclohexane
http://www.atsdr.cdc.gov/toxprofiles/tp43-c1.pdf
gespeichert am: 01.11.2007
Baumann, K., Angerer, G., Lehnert, G., Brassow, H.-L. (1983): Chronische Belastung
des Menschen mit Hexachlorcyclohexan. In: DFG (Hrsg.), Hexachlorcyclohexan als
Schadstoff in Lebensmitteln. Verlag Chemie Weinheim
Braun,R., Fuhrmann (1999): Spezielle Toxikologie für Chemiker. Eine Auswahl
toxischer Substanzen. Teubner Studienbücher Chemie, Stuttgart; Leipzig
Daunderer, M. (1991): Umweltgifte. Diagnostik und Therapie. Ecomed, Lanfsberg/ Lech
Dekant, W.; Vamvakas, S. (1994): Toxikologie für Chemiker und Biologen. Spektrum,
Akad. Verl. Heidelberg; Berlin; Oxford, 290-291
DFG (1982): Deutsche Forschungsgemeinschaft (Hrsg.). Hexachlorcyclohexan-
Kontamination – Ursachen, Situation und Bewertung, Kommission zur Prüfung von
Rückständen in Lebensmitteln, Mitteilung IX. Harald Boldt Verlag, Boppard.
Eichler, D. (1983): Physikochemische Eigenschaften, Verhalten und Analytik der HCH-
Isomeren. In: DFG (Hrsg.), Hexachlorcyclohexan als Schadstoff in Lebensmitteln. Verlag
Chemie Weinheim
Eichler, D. (1983): Biotisches und abiotisches Ab- und Umbauverhalten einschließlich
Isomerisierung (Pflanze). In: DFG (Hrsg.), Hexachlorcyclohexan als Schadstoff in
Lebensmitteln. Verlag Chemie Weinheim
Engst, R., Macholz, R.M., Kujawa, M. (1977): Recent state of lindane metabolism. In:
Residual: Reviews Residues and other contaminants in the total environment. Vol. 68,
Springer-Verl., New York; Heidelberg; Berlin
EPA (2000): www.epa.gov/ttn/atw/hlthef/lindane.html,
gespeichert am 30.10.2007
Page 34
Literaturverzeichnis
34
Fabre, B.; Roth, E.; Kergaravat, O. (2005): Analysis of the insecticide
hexachlorcyclohexane isomers in biological media. A review. Environ. Chem. Lett. 3, 122-
126
Falbe, J.; Regitz, M. (Hrsg.) (1997): Roempp Chemielexikon. Georg Thieme Verlag,
Stuttgart; New York
Gao, H.J.; Jiang, X.; Wang, F.; Wang, D. Z.; Bian, Y. R. (2005): Residual Levels and
Bioaccumulation of Chlorinated Persistent Organic Pollutants (POPs) in Vegetables from
Suburb of Nanjing, People’s Republic of China. Bull. Environ. Contam. Toxicol. 74, 673-
680
Gonzalez, M.; Miglioranza, K.S.B.; Aizpun de Moreno, J.E.; Moreno, V. (2003): Occurence and Distribution of Organochlorine Pesticides (OCP) in Tomato (Lycopersicon
esculentum) Crops from Organic Production. J. Agric. Food Chem. , 51, 1353-1359
Haider, K. (1983): Abbau und Umwandlung von γ-HCH und anderen HCH-Isomeren
durch Bodenmikroorgansimen. In: DFG (Hrsg.), Hexachlorcyclohexan als Schadstoff in
Lebensmitteln. Verlag Chemie Weinheim
Heinrich, K. (1997): Untersuchungen zum Boden-Pflanze-Transfer ausgewählter
organischer Umweltschadstoffe in Abhängigkeit von Bodeneigenschaften. UFZ-Bericht
Nr. 11/1998. ISSN 0948-9452.
Herbst, M.; Bodenstein, G. (1973): Toxikologie des Lindans. In: Ulmann, E. (Hrsg.):
Lindan. Monographie eines insektiziden Wirkstoffs. Verlag K. Schillinger, Freiburg im
Breisgau
Herbst, M. (1983): Toxizität von γ – HCH (Lindan) In: DFG (Hrsg.), Hexachlorcyclohexan
als Schadstoff in Lebensmitteln. Verlag Chemie, Weinheim
Hoffmann, A. (1983): Toxizität von α- und β-HCH. In: DFG (Hrsg.), Hexachlorcyclohexan
als Schadstoff in Lebensmitteln. Verlag Chemie, Weinheim
Jensen, A.A. (1983): Chemical contaminants in human milk. Res. Rev. 89, 1-128
Kawahara, T., Nakamura, H. (1971): Organochlorine pesticide residues in plants and
soils. Absorption and translocation of organochlorine pesticides in tobacco. Nayaku
Kenshasho Hokoku 11, 61
Page 35
Literaturverzeichnis
35
Koch, R. (1986): Datenspeicher Umweltchemikalien. Eigenschaften, Toxizitäten,
Umweltverhalten. VEB Verlag Volk und Gesundheit Berlin
Korte, F. (1983): Verhalten von HCH-Isomeren im Boden. In: DFG (Hrsg.),
Hexachlorcyclohexan als Schadstoff in Lebensmitteln. Verlag Chemie Weinheim
Korte, F. (1992): Lehrbuch der Ökologischen Chemie. Grundlagen und Konzepte für die
ökologische Beurteilung von Chemikalien. Georg Thieme Verlag, Stuttgart; New York
Landesanstalt für Umweltschutz Baden-Württemberg (LfU) (1998): Literaturstudie
zum Transfer von organischen Schadstoffen im System Boden/ Pflanze und Boden/
Sickerwasser.
Landesanstalt für Umweltschutz Baden-Württemberg (1993): Stoffbericht
Hexachlorcyclohexan (HCH). Texte und Berichte zur Altlastenbearbeitung
Li, H.; Sheng, G.; Sheng, W.; Xu, O. (2002): Uptake of trifluralin and lindane from water
by ryegrass. Chemosphere 48, 335-341
Marquardt, H.; Schäfer, S.G. (Hrsg.) (1994): Lehrbuch der Toxikologie. BI-
Wissenschaftsverlag, Mannheim; Leipzig; Wien; Zürich
Pawliszyn, J.(1997): Solid Phase Microextraction. Theory and Practice. Wiley-VCH, New
York; Chichester; Weinheim; Brisbane; Singapore; Toronto
Portig, J., Vohland, H.W. (1983): Neuropharmakologische und neurotoxische Wirkungen
von Hexachlorcyclohexan (HCH), In: DFG: Hexachlorcyclohexan als Schadstoff in
Lebensmitteln. Verlag Chemie Weinheim
Scheffer,F.; Schachtschabel, P. (2002): Lehrbuch der Bodenkunde. Spektrum Akad.
Verlag, 15. Auflage, Heidelberg; Berlin
Schulte-Herrmann, R. (1983): Zum Mechanismus der Tumorbildung in der Leber unter
dem Einfluss von HCH-Isomeren. In: DFG: Hexachlorcyclohexan als Schadstoff in
Lebensmitteln. Verlag Chemie Weinheim
Streit, B. (1991): Lexikon Ökotoxikologie. VHC, Weinheim
Tao, S.; Xu, F.L.; Wang, X. J.; Liu, W. X.; Gong, Z. M.; Fang, J.Y.; Zhu, L. Z.; Luo, Y. M. (2005): Organochlorine Pesticides in Agricultural Soil and Vegetables from Tianjin,
China. Environ. Sci. Technol. 39, 2494-2499
Page 36
Literaturverzeichnis
36
Umweltbundesamt (UBA) (2006): Mulde-Fische weiter mit Hexachlocyclohexan (HCH)
belastet. Hintergrundpapier
www.umweltbundesamt.de/uba-info-presse/hintergrund/HCH_in_Elbefischen.pdf
gespeichert am: 30.10.2007
Verma, A. und Pillai, M. K. K. (1991): Bioavailability of soil-bound residues of DDT and
HCH to certain plants. Soil Biol. Biochem., Vol. 23, No. 4, 347-351
Vohland, H.-W., Koransky, W. (1983): Zu Verhalten und Wirkung des
Hexachlorcyclohexans beim Menschen. In: DFG (Hrsg.), Hexachlorcyclohexan als
Schadstoff in Lebensmitteln. Verlag Chemie, Weinheim
Willett, K.L., Ulrich, E.M., Hites, R.A. (1998): Differential toxicity and environmental
fates of Hexachlorocyclohexane isomers. Environmental Science and Technology, Vol.
32, No. 15, 2197-2207
WHO (1998): Selected non-heterocyclic polycyclic aromatic hydrocarbons, Geneva,
World Health Oragnization, International Programme on Chemical Safety, Environmental
Health Criteria 202
http://www.inchem.org/documents/ehc/ehc/ehc202.htm
gespeichert am 12.07.2006
WHO (1991): Environmental Health criteria 124: Lindane (Gamma-HCH), Health and
Saftey Guide, Geneva
http://www.inchem.org/documents/hsg/hsg/hsg054.htm
gespeichert am: 12.07.2006
WHO (2003): Health risks of Persistent Organic Pollutants from long-range
transboundary air pollution
http://www.lfu.bayern.de/umweltwissen/schadstoffe/doc/stoffbeschreibungen/12_lindan.p
df
gespeichert am: 30.10.2007
Rückstands-Höchstmengenverordnung (RHmV): 1999
Page 37
Tabellen- und Abbildungsverzeichnis
37
Tabellenverzeichnis Tabelle 1: Physikalische Eigenschaften von α-, β- und γ-HCH …………………..6
Tabelle 2: HCH-Vergiftung der Ratte: Neurotoxische Zustandsbilder ………………15
Tabelle 3: Trennbedingungen bei den GC/MS Analysen …………………………….26
Tabelle 4: Lindan-Konzentrationen zu Versuchsbeginn sowie Rückstände in
Boden und Nährlösung nach einer Vegetationsperiode ………………...27
Tabelle 5: Lindan-Konzentrationen in den Pflanzen …………………………………28
Tabelle 6: Gesamtgehalte und Aufnahmeraten ………………………………………29
Tabelle 7: Ergebnisse der in vivo – Messung ………………………………………..29
Tabelle 8: Transferfaktoren Boden-Pflanze (Tomate) von γ-HCH ………………….30
Abbildungsverzeichnis
Abbildung 1: Darstellung des HCH in der Sesselform …………………………………...4
Abbildung 2: Stereoisomerie des HCH, axiale und äquatoriale Position der Cl-Atome . 5
Abbildung 3: Möglicher Abbauweg von γ-Pentachlorcyclohexen-1 in höheren
Pflanze ……………………………………………………………….12
Abbildung 4: Metabolismus von Lindan bei Säugetieren ……………………………..14
Abbildung 5: Wirkungsmechanismus der insektiziden Organochlorverbindungen ….. 16
Abbildung 6: SPME – Faser ………………………………………………………………..21
Abbildung 7: Auswahl des Fasermaterials ………………………………………………22
Abbildung 8: Kalibrationsgerade für die Bestimmung von Lindan mittels SPME aus
Wasser …………………………………………………………….23
Abbildung 9: Anreicherung des Lindans aus der Fruchtmatrix mittels SPME ……… ..25
Abbildung 10: Matrixabhängigkeit ………………………………………………………25
Abbildung 11: Lindan-Konzentrationen in den Pflanzenproben …………………………28
Abbildung 12: SPME – Faser im Stamm einer Tomatenpflanze zur in vivo – Messung 30
Page 38
Abkürzungsverzeichnis
38
Abkürzungsverzeichnis Abb. Abbildung
BRD Bundesrepublik Deutschland
bzw. beziehungsweise
DFG Deutsche Forschungsgemeinschaft
d.h. das heißt
DNA Desoxyribonukleinsäure
EPA Environmental Protection Agency
GABA γ - Aminobuttersäure
GC Gaschromatographie
HCH Hexachlorcyclohexan
Hrsg. Herausgeber
KG Körpergewicht
LD50 mittlere tödliche Dosis 50
LfU Landesanstalt für Umweltschutz
KOW n-Oktanol/Wasser-Verteilungkoeffizient
MS Massenspektrometrie
NOAEL no observed adverse effect level
PA Polyacrylat
PDMS Polydimethysiloxan
PDMS/DVB Polydimethysiloxan/ Divenylbenzen
RHmV Rückstands - Höchstmengenverordnung
SPME Festphasenmikroextraktion (Solid Phase Microextration)
Tab. Tabelle
TF Transferfaktor
TG Trockengewicht
u. a. unter anderem
UBA Umweltbundesamt
U/min Umdrehungen pro Minute
v. a. vor allem
WHO Weltgesundheitsorganisation (World Health Organisation)
z. B. zum Beispiel
ZNS Zentrales Nervensystem
Page 39
39
Eidesstattliche Erklärung Hiermit versichere ich, dass ich die vorliegende Arbeit ohne unzulässige Hilfe Dritter und
ohne Benutzung anderer als der angegebenen Hilfsmittel angefertigt habe.
Nadine Zeiner
Leipzig, 12.12.2007